- Existence of neutrality and near-neutralityPart II-Neutral evolution is relevant
-Evolution lacks foresight; it can neither anticipate nor respond-Clarifying some terminology: two types of function, positive vs. negative selection-Rise of complexity through non-adaptive means
-An example of constructive neutral evolution at work: loss of group I intron self-splicingPart III
-Further examples of constructive neutral evolution
-Discussion of what sparked this argument: Evolution of ciliate nuclear dimorphism
Neutral evolution is relevantAgain, apologies for stating the obvious, but apparently even some prominent
evolutionary biologists popularisers of evolutionary biology fail to grasp this simple concept. I've masticated this point to a fine mush by now, but selection and neutral processes
act in tandem. Mutational bias and drift
matter. So often in the literature you find people arguing over whether something is an adaptation or a spandrel. This gets even more absurd when the structure in question is as massive as the
human language capacity. For example, in the landmark
Pinker & Bloom 1990 paper signalling the revival of evolutionary linguistics, you find awful sentences like:
"The key point that blunts the Gould and Lewontin critique of adaptationism is that natural selection is the only scientific explanation of adaptive complexity." [p.6; emphasis mine]
First off, it's kind of cute that a couple psychologists seem to think they can so easily outright dismiss a point made by evolutionary biologists. I mean, seriously, I find it adorable. On that note, I'm now gonna write a book chapter debunking generative linguistics, because, well, my buddy says they're wrong. I wonder how much Pinker's evolutionary views have been shaped by Dawkins
et al. Their camp is rather influential outside evolutionary biology, and while many claim that panadaptationism is a strawman -- and even in biology that point is debatable -- outside evolutionary biology, panadaptationism is alive and well. Part of the reason is that adaptationist stories are written in popular books, while pluralistic approaches largely remain hidden in the likes of
Molecular Biology & Evolution,
Biology Direct and
Journal of Molecular Evolution. Researchers working in applied evolutionary fields have likely never heard of them.
Back to Pinker & Bloom, first off it's quite a tautology to claim that adaptive complexity evolves through adaptation. Well, yes, you JUST labelled it adaptive. In the immortal words of 4chan, long cat is loooooong. Casting that aside, they've falled for a false dichotomy. You would think someone as smart as Steven Pinker and Paul Bloom wouldn't fall for it. But they did. Why does it have to be one
or the other? Why adaptation
or spandrel? Especially when we speak of highly complex systems -- is it not in the definition of complexity that they consist of multiple components? How likely is it that
all of them are adaptive or neutral or maladaptive? Is it even remotely productive to reduce a system so drastically as to label it simply as an adaptation? What does that even mean, besides stating the obvious? 'Adaptation' has got to be one of the more utterly
useless terms in evolution biology, at least the way it's abused today.
Letting Pinker & Bloom speak further:
"Adaptive complexity" describes any system composed of many interacting parts where the details of the parts' structure and arrangement suggest design to fulfill some function. The vertebrate eye is the classic example. The eye has a transparent refracting outer cover, a variable-focus lens, a light-sensitive layer of neural tissue lying at the focal plane of the lens, a diaphragm whose diameter changes with illumination level, muscles that move it in precise conjunction and convergence with those of the other eye, and elaborate neural circuits that respond to patterns defining edges, colors, motion, and stereoscopic disparity. It is impossible to make sense of the structure of the eye without noting that it appears as if it was designed for the purpose of seeing -- if for no other reason that the man-made tool for image formation, the camera, displays an uncanny resemblance to the eye. Before Darwin, theologians, notably William Paley, pointed to its exquisite design as evidence for the existence of a divine designer. Darwin showed how such "organs of extreme perfection and complication" could arise from the purely physical process of natural selection." [p.6 cont'd]
Holy fucking crap,
argument from design,
for selectionism! Impressive. Yes, you guys just totally pwned Gould with the vertebrate eye. He was unaware of its very existence. Eyes don't preserve well in the fossil record, you see? Eyes look designed, therefore selection. Great. I'll let them finish...
"The essential point is that no physical process other than natural selection can explain the evolution of an organ like the eye." [p.6]
AFAIK, Gould never
denied selection!!! He was a brilliant biologist who thoroughly understood evolution, unlike some
recent drama queens. What Gould argues is that
a) not everything is an adaptation and
b) adaptation is neither the sole nor the most important 'force' in evolution (nor is it actually a 'force' of any sort...). Furthermore, while I am unaware of Gould's opinions about The Eye, and am currently too lazy to research the topic, to me it seems highly implausible that the vertebrate eye evolved solely through selection. In fact, considering that selection is a
purifying, not driving, 'force' -- that is, selection simply removes the not sufficiently fit -- it is curious to see where Pinker & Bloom think the 'material' for the selective evolution of the eyes comes from. Surely they're not insane enough to believe that the thing evolved entirely through point mutations each making the eye progressively slightly better and better and suddenly, veeeery gradually, ta-da: The Eye!
On that note, I wonder if there's a strong link between selectionism and gradualism. In a sense, one does kind of have to believe the above point-by-point scenario to explain how anything arises
purely by selection. To anyone with the slightest inkling of how genes and genomes work, such a view is obviously absurd. Although considering how Dawkins has already
enlightened us that molecular biologists may or may not be -reputable- biologists...
I'm still amused by how remarkably cute it is of Pinker & Bloom to know so much about the details of evolution that they can make bold statements like the last sentence cited above. That's one
strong statement!
tl;dr Some rather reputable and smart people still fall for the false dichotomy where something is either purely adaptive or purely 'random'. Selection is
not the sole source of order (
Lynch 2007 PNAS; you might as well head over and read that paper by now)
So if selection is not the only source of order, what else could be? How can neutral forces (coupled with negative selection) result in an increased complexity? Ah, nearly time for constructive neutral evolution. But prior to that, one more thing to take care of...
Evolution lacks foresight; evolution is not engineeringFirst of all, one must note that a modern function of any given system may not necessarily have been there at the initial stages of evolution. In fact, it is often counterproductive to even assume so. A nice example would be diatom sex.
Upon each division, one of the diatoms gets smaller. The cell is surrounded by two valves: a larger top valve and a smaller bottom one. During division, the bottom one actually becomes the top in the daughter cell. After several generations, some diatom offspring get a little on the small side. Luckily, sex makes them bigger; ie. they produce gametes (or swap nuclei), fuse and make a brand new maximum-sized cell. Jennifer Frazer
wrote an awesome post on this topic.
One may look at diatoms and say: What is the function of sex? If you remove sex, the diatoms can't get bigger again, and die a miniscule death. Thus, sex must be there to get bigger!
However, one must imagine evolution as a blind step-by-step process, devoid of any foresight or momentum whatsoever. A common fallacy would be to think of the diatoms first having a problem, and then having to come up with an emergency solution to it, eg. evolve sex. This may seem obviously ridiculous in the case of something as complex as sex, but is often assumed for simpler cases, like evolving a resistance to something. Perhaps we get this picture from the antibiotic resistance examples in bacteria; however, antibiotic resistance is generally much too simple, and bacteria much too numerous, to apply this mode of evolution to everything else. It is additionally pretty rare, but we don't notice the vastly overwhelming multitude of unsuccessfulcases.
Back to our diatoms, the more plausible scenario is actually that sex was there first, and was able to correct for the problems caused by this rather suboptimal mode of mitosis. Sex
enabled this peculiar life style; it did not evolve in response to it. 'Enable' is a word that must be used much more frequently in the literature. Certain situations
enable certain mutations or other changes to persist or excalate into fixation (either by drift or selection). They do not actually push them there.
Again, evolution cannot anticipate.
Two meanings of 'function'As a brief aside, it is important to note two distinct meanings of the term 'function', often used rather carelessly:
1.
Selected function - what a certain trait would have been selected for in its evolutionary past.
2.
Current function - based upon what would happen were the trait removed
in the present.
Going back to our diatom example above, the current function of sex in diatoms can be argued as a way to increase the population size back to original levels; for inhibiting sex would remove a way for the organisms to get bigger again. However, this is not the selected function of sex, since it must have already existed before the size problem happened. (the selected function of sex, if any, remains a fuzzy, murky mess)
Positive vs. negative selectionOne more brief thing to clean up: 'types' of selection
Negative (aka purifying) selection - selection acting
against a trait
Positive selection - selection
promoting a trait in a population, usually through competition (the type most commonly spoken of in popular media, despite being far less common to the point of relative rarity...)
Ultimately, positive selection is a form of negative selection where the appearance of a fitter form of a trait renders the rest of the variants relatively less fit, resulting in them being selected
against (negative selection). Thus, selection isn't actually ever
for anything, strictly speaking. Selection is a constraint, weeding out variants that are insufficiently fit. Furthermore, selection is always lurking about in background, albeit in varying intensities depending on the conditions (
see Part I). That said, one must not ever conflate selection with adaptation -- they are
not the same thing by any means!
Rise of complexity through non-adaptive meansI won't go into much detail here as I'm sure Arlin Stoltzfus himself will explain it much better in
his upcoming posts on Sandwalk. Also, unlike me, he's actually qualified to write about this stuff. I will give a crude overview (to the best of my understanding) along with a few concrete examples. For an additional source on this, I blogged
Ford Doolittle's seminar talk on the subject here. The non-adaptive evolution of complexity requires tinkering, relaxed selection (eg. due to smaller effective population size) and ratchets. Tinkering is basic (as mentioned above, evolution lacks foresight, many adaptations are far from optimal as relics of past functions are retained, etc) so I won't discuss it. Small population sizes were discussed in
Part I, so now we need the final piece: Ratchets. This is where Constructive Neutral Evolution enters.
In a nutshell, chance interactions may happen to compensate for otherwise-deleterious mutations, thereby enabling them to eventually occur, thereby ultimately resulting in an irreversible dependency upon the interaction. That way, complexity (loosely defined as number of components and interactions within a specified system; as in
Lukes et al 2009 PNAS) can increase without ever actually being adaptive. This mechanism stresses that not all complexity is actually 'better', and perhaps much of it may well be a case of mere bloating, much like the bureaucracy of an institution with time. For a system that is required to be constitutively functional, it is in fact easier to bloat complexity than to trim it down, partly just due to basic combinatorics: there are many more ways of being complicated than being simple, and just statistically there'd be more viable states of higher complexity.
 Overview of Constructive Neutral Evolution. Modified and enhanced from older diagram here; Ford Doolittle uses something similar in his talk, so it shouldn't be too far off.
Overview of Constructive Neutral Evolution. Modified and enhanced from older diagram here; Ford Doolittle uses something similar in his talk, so it shouldn't be too far off.This model, while being employed more in molecular biology, is actually applicable universally. Ratchets occur in ecology (someone brought up ecological specialisation as an example), sociology (your favourite institutional bureaucracy), and even evolutionary linguistics (eg. many types of change are irreversable or unlikely to be reversed). Going back to our non-anticipatory step-by-step evolutionary thinking: constructive neutral evolution is
powerful. It provides the 'push' where adaptation cannot explain a particular increase in complexity. Ratchets provide an appearance of directionality to a system.
Cyt-18 and the N.crassa mitochondrial self-splicing intronI will discuss the not-so-self-splicing intron examples in further detail in part III, but just to provide a quite example to the above point, let's look at a
Neurospora mitochondrial self-splicing intron that developed a dependency on another protein. This example is based on an old-ish paper,
Akins & Lambowitz 1987 Cell. It was found that mutants of
cyt-18 exhibited defects in the splicing of group I (self-splicing) introns. Curiously,
cyt-18 turned out to be homologous to tRNA synthetases. Further work confirmed that
cyt-18 is necessary for splicing in
N.crassa, and phylogenies showed it to be a derived trait. That is, in closely related lineages,
cyt-18 is NOT necessary for group I intron splicing.
Now the question is, why would N.crassa require an additional protein to perform the same process its relatives can do without involving extra bits. The knee-jerk adaptationist reaction would be to think of it as being somehow better that the simple fully-self-splicing variant. Perhaps the use of a protein enables a better regulation of splicing. For some reason, any peculiar complexity in genetics has to be there for regulation. Apparently, evolution is directionless but still strives for higher complexity of regulatory systems, if you listen to molecular geneticists. However, is this necessarily true for all instances of higher complexity of interactions? How else could complex interactions arise if not for the improvement of a system?
This is where our friendly neighbourhood ratchet, constructive neutral evolution, comes in. Suppose initially the group I intron spliced itself out without the help of any proteins. Just incidentally, a random protein, in this case cyt-18, just happened to bind to it at times. Anyone who's worked with yeast-two-hybrid systems or done immunoprecipitation would be painfully aware of 'false' interactions; that is, when proteins interact biochemically but without any biological significance; or very rarely bind in vivo to begin with. The network of such fortuitous interactions is vast, and increases rapidly as more components are involved -- a positive feedback loop.
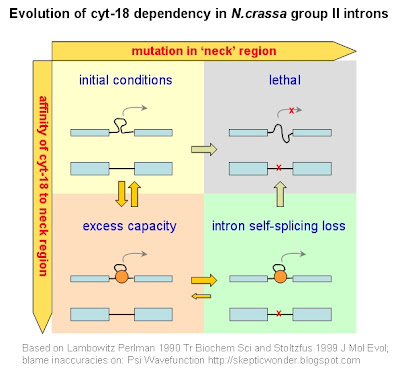
In the Akins & Lambowitz example, the self-splicing intron needs to form a particular secondary RNA structure to remove itself. For the sake of simplicity, let's pretend it has a single neck leading to a loop. The neck needs to form properly in order for the splicing to work, so any mutation destabilising the neck would be lethal. This means these mutations are 'impossible' in the evolutionary sense, as we're unlikely to ever see them.
Now let's say this cyt-18 protein happens to bind to the neck region, and stabilise it slightly. In the grand scheme of things, the intron can splice out properly regardless of whether this protein is there, so it doesn't increase fitness in any way. However, what this protein does to is create an excess capacity in the system by enabling certain otherwise-deleterious mutations to arise in the neck region by compensating their destabilisation effects. Once such a mutation occurs (and it's bound to happen eventually), the lineage now depends upon cyt-18 for proper splicing to occur. Since the chance of the mutation being reverse is quite small, and often even less than the chance of another otherwise-deleterious mutation further fixing the protein dependency, the system is now stuck depending on cyt-18, with no adaptive advantage whatsoever. In a diagram:
N.crassa mitochondrial self-splicing intron example of constructive neutral evolution. See text. Based on Akins & Lambowitz 1987 Cell, Lambowitz & Perlman 1990 Tr Biochem Sci, Stoltzfus 1999 J Mol Evol.
In such a manner, interactions can rapidly accumulate and form the ridiculously intricate pathways that horrify biochemists, geneticists and cell biologists alike, not to mention anyone working with 'transcriptomes' and 'proteomes' and (ewww) 'interactomes'. Not all of these complex pathways are that way for adaptive reasons, despite the common assumption. Some of these complexities can later be exapted for adaptive reasons, sure, but arguably most didn't initially arise adaptively. See
Lynch 2007 Nat Rev Genet for a review on neutral evolution of gene interaction pathways.
My hunch at the moment, which may or may not have anything to do with reality, is that most complexity initially may arise through constructive neutral evolution, and some of it later drifts into doing something useful, upon which the lineage fixes a dependency, occasionally through 'positive' selection. Neutral explanations for complexity alleviate the need for awkward scenarios where an organism desperately needs to evolve something quickly as a response. The chance that the right features evolve at the right time, and not before the whole lineage dies off, is usually not very high. But when a lineage has been 'preadapted' through such neutral means, it wouldn't be as adversely affected by changes, or by finding a new niche, or whatever. Evolutionary adaptation is not a particularly efficient response mechanism, but neutral evolution is a wonderful source of 'enabling' modifications (excess capacities) that can later be molded into apparently functional parts. The explosion of complexity is controlled by negative selection, preventing it from getting too extreme and costly, although in organisms with smaller effective population sizes (where drift often overpowers selection; see Part I), complexity can get quite extreme nonetheless.
Most supporters of neutral theories and pluralism perhaps wouldn't be as extreme though, so don't take the above paragraph too seriously. In Part III, I will discuss further examples, including the little that is known about the evolution of ciliate nuclear dimorphism as well as gene scrambling, which was the original topic that sparked this discussion. Furthermore, I'd like to try and extrapolate this model beyond molecular biology, venturing a bit into evolutionary linguistics, which may also be teeming with ratchets. Hopefully by now you'd see that the concept of neutral evolution being capable of driving complexity is at the very least worth considering.
As Michael Lynch pointed out (eg.
2007 PNAS), selection may very well not be the sole source of order and complexity. In some cases, perhaps not even the dominant one.
ReferencesAkins, R., & Lambowitz, A. (1987). A protein required for splicing group I introns in Neurospora mitochondria is mitochondrial tyrosyl-tRNA synthetase or a derivative thereof Cell, 50 (3), 331-345 DOI: 10.1016/0092-8674(87)90488-0
LAMBOWITZ, A., & PERLMAN, P. (1990). Involvement of aminoacyl-tRNA synthetases and other proteins in group I and group II intron splicing Trends in Biochemical Sciences, 15 (11), 440-444 DOI: 10.1016/0968-0004(90)90283-H
Lukes J, Leander BS, & Keeling PJ (2009). Cascades of convergent evolution: the corresponding evolutionary histories of euglenozoans and dinoflagellates. Proceedings of the National Academy of Sciences of the United States of America, 106 Suppl 1, 9963-70 PMID: 19528647
Lynch, M. (2007). Colloquium Papers: The frailty of adaptive hypotheses for the origins of organismal complexity Proceedings of the National Academy of Sciences, 104 (suppl_1), 8597-8604 DOI: 10.1073/pnas.0702207104
Lynch, M. (2007). The evolution of genetic networks by non-adaptive processes Nature Reviews Genetics, 8 (10), 803-813 DOI: 10.1038/nrg2192
Pinker S, & Bloom P (1990). Natural language and natural selection Behavioral and brain sciences, 13 (4), 707-784
Stoltzfus, A. (1999). On the Possibility of Constructive Neutral Evolution Journal of Molecular Evolution, 49 (2), 169-181 DOI: 10.1007/PL00006540 


















































{kind=link}
{kind=link}