
Still procrastinating with the
Heterolobosea posts (no, I haven't forgotten). Real protistologists give me sorry looks when I mention being stuck writing about that rather obscure and messy group. I don't want to just do a taxonomic overview - I like to mention odds and ends about their cell biology as well. Sadly, the cell biology of most of those things is a sorry neglected mess. Even the eruptive pseudopodia that are quite characteristic of this group (although present in some amoebozoans as well) seem to not have been examined on a molecular level - I cannot find a single paper describing the cytoskeletal dynamics behind this peculiar mode of motility, and my sources point out a severe shortage of attention in that area. But I'm still scraping things together, so
eventually something will appear...
Now for something completely different...since someone just sent me scouring piles of archaic
metazoan '
mesozoan' literature to identify
that damn bug (an orthonectid of some sort), I have a random metazoan of my own to show off a bit, from the latest pond microforay:


Rotifer. (Mine =P 40x* DIC) Note the contractile vacuole-like structure (actually its bladder, but appears to function in a very similar way. Another case of ultimate convergence?).I can't quite key this one out, especially since this damn sucker is on its side. Probably the only reason I was able to capture a photo of it, since they tend to be quite hyperactive normally. This one appears to be immobilised by the cover glass. Perhaps it may be
Aspelta or something from the
Lepadellidae. Or might it be a Bdelloid? Perhaps a certain taxonomist could help out. Especially the one who made me browse orthonectids for the past few hours. ^^
*40x here, and onwards, refers to objective rather than magnification. We don't actually care about mag, it's the other properties of the optical system that matter much more when dealing with professional instruments. (Such as numerical aperture and field of view, focal length, focal depth, which type of immersion medium its calibrated for, etc) Technically, I should be writing down the NA and such, but we're not that meticulous crazy around here.
**Randomly found out there's a rotifer that lives inside colonies of Volvox, as a parasite...Much of my resentment towards metazoa simply comes from them being overstudied and overemphasised,
to the neglect of other phyla. That said, they are still interesting and still poorly understood lifeforms, especially among the spineless things. While representing but a mere sliver of life's overall diversity -- contrary to what our senses tell us -- metazoa are nevertheless, like all life, fascinating and
weird. Metazoan multicellularity, as mundane and ubiquitous as it may seem to our rather biased eyes, is about as weird and insane as kinetoplastid kDNA editing, or the convoluted ciliate nuclear genomes, or the breathtakingly massive, and seemingly pointlessly so, nuclear genomes of dinoflagellates and some amoebozoa. I'm quite disappointed at the absense of the 'perspective of strangeness', if you will, that seems to dominate many animal-oriented classes (especially those catering to pre-meds. Must. not. rant.)
Interestingly, off all taxonomic and phylogenetic literature, I find metazoan papers to be the most difficult to follow* - we actually have common names to refer to quite a few
animals metazoa. Furthermore, the obsession with ranked hierarchy is driving me insane - you DO NOT have to name every single possible grouping you can come up with, seriously! That makes searching for stuff an absolute disaster, as you have to look for the relevant subfamily, family, superfamily, suborder, order, superorder, hypersuperorder, subclass... etc. All named differently enough to not fit into one convenient Google (or PubMed, or WebofScience) search. And you have to find all of them. And they change. And your life becomes a living hell. Please stop, seriously! WHYYYYY!? Did TC-S write their taxonomies or something?! And they listened to ALL of it?! [/rant]
Another fascinating facet of biodiversity that seems to be thoroughly ignored is the interface between kindoms: the breathtaking diversity and strangeness of the possible relationships between the many corners of life. Of course we get the 'animals eat plants' story**, occasionally hear of animal-earing plants. Then we hear of fungi and bacteria as decomposers. If you hang out on the botany side of the Great Divide, you also hear of algae as the aquatic producers and fungi as mycorrhyzal symbionts (plant-fungal relationships are a fascinating world of awesome, by the way). But that's pretty much all ecology you ever get (outside of ecology programs anyways. I hope!), and frankly, it's pretty boring.
What about the vast 'multicultural' realm of the customs and ways of the parasites? What about the unexpected mutual endosymbioses, both of multicellular and unicellular things, on both multicellular and unicellular levels? What about multicellular predators, and parasites(!) of unicellular organisms? Or unicellular predators(!) of multicellular organisms? For a case of the latter, let's turn back to our rotifers. By the way, if you're desperate for a tree, or sexy SEMs of rotifer morphological features,
this paper could come in handy(free access).
*Well, that and any prokaryotic literature. Bacteria are...complicated. Unlike Eukaryotes. ^_^**A while ago while writing a biochem exam, I came across a question starting off with plant lipid synthesis. I got excited (OMG they're mentioning PLANTS!!!). Then I read on: "...which is then consumed by a herbivore." and wanted to throw something at whoever wrote that question. That pretty much sums up the biochemists' and zoologists' view of plants.Quick aside: the border between parasite and predator is quite fuzzy and arbitrary, especially once you venture outside familiar grounds. Generally, people tend to call something a parasite when it feeds of something bigger than itself, and a predator when it feeds off something smaller.
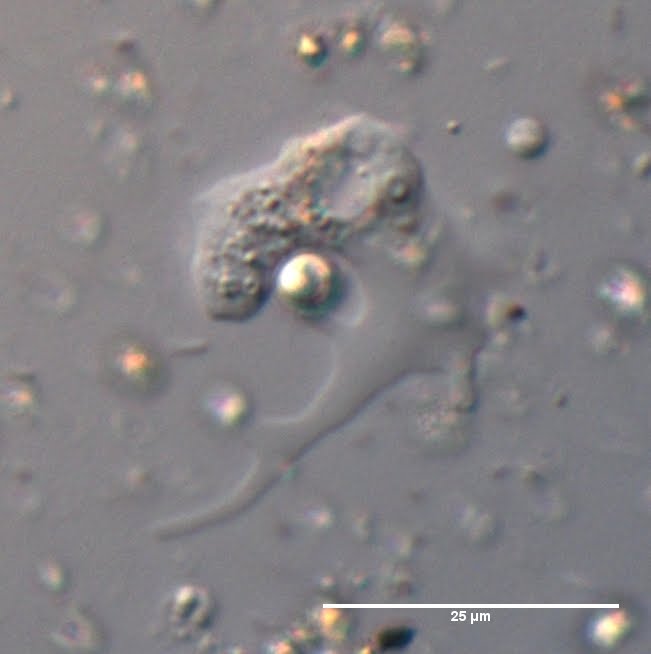
Rotifers, like anything else out there, have no choice but to participate in the great web of things eating things. As is the case for many small pond organisms, their predators are plentiful. In fact, there's some rather embarassing ones. Including a relative of this slow thing:
(Mine again. Having my own photo stock would save me so much search and citation time. Of course, making my own stock might kind of negate that.)
Yeah, rotifers can get chomped on
by Difflugia, of all things.
The [planktonic thecate(!)] amoeba senses the
thecate rotifer,
quickly extends its pseudopodium along its length as if to 'measure' it, positions itself with its 'mouth' towards the rotifer's foot, somehow makes a hole in its protective jelly, and slurps in the rotifer's contents, while grasping the shell of its prey with the pseudopodia. All within 20min. Amazingly, this amoeba is not a rotifer specialist - it only eats them upon a rare chance encounter. Thus, the amoeba actually has complex behavioural patterns, enabling it to detect the type of prey, sense its shape and decide on an attack strategy.
Many protist predators are actually quite picky, sifting through the various detritus they come across and deciding on what to engulf. Combine that with eyespots and ocelli (Warnowiid dino camera eyes!), along with other oddities of the unicellular world, and we've got ourselves a whole field of Protist Behavioural Biology.
As we look more and more carefully at the unicellular world, it becomes increasingly more apparent that cells can 'think' -- not in the woo-ey "OMG feelings and consciousness!!1!" kind of way (in fact, I HATE that shit) -- but in terms of processing information from their environment in a rather complex way. Doesn't seem like much is known about cellular 'molecular intelligence' (or 'molecular instinct' - the line between canonical behaviour and genetic/biochemical pathways is quite blurry, it seems), and the field is dominated by junk thinking at the moment. And computer scientists, who don't seem to be bothered by biological reality. But hopefully as more and more results pile up suggesting some sort of 'cellular intelligence*', some fundamentally interesting stuff may come out of that research.
*I feel I may end up hating myself for using that term, but I can't think of anything better at the moment...sadly, too many good words get hijacked by sloppy thinking.If you ever find yourself randly shrinked to the micron scale some day: Stay the fuck AWAY from amoebae. They may look slow and stupid and oozy, but they're out to fucking get you. They vicious. They don't care what unicellular or multicellular phylum you happen to be a proud member of: if they can hug you, they WILL squish you with nasty enzymes. Or make holes in you. They're freaking SCARY! o_O
Same goes for forams and "radiolaria", by the way. Actually, on the micron scale, if something doesn't eat you from the outside, chances are, it's patiently waiting to devour you from the inside. The microbial world tends to be quite innovative. And we haven't even gotten to the
lethal veil of pallium...
Starting to feel good about your size yet?
As much as I'm obsessed with the microscopic world, it feels much safer to be a mere observer, rather than a participant. There's some truly terrifying monsters in those waters!
ReferencesHan, B., Wang, T., Lin, Q., & Dumont, H. (2007). Carnivory and active hunting by the planktonic testate amoeba Difflugia tuberspinifera Hydrobiologia, 596 (1), 197-201 DOI: 10.1007/s10750-007-9096-zRiemann, O., Kieneke, A., & Ahlrichs, W. (2009). Phylogeny of Dicranophoridae (Rotifera: Monogononta) - a maximum parsimony analysis based on morphological characters Journal of Zoological Systematics and Evolutionary Research, 47 (1), 61-76 DOI: 10.1111/j.1439-0469.2008.00482.x




























































{kind=link}