Was forwarded a great article on similarities between creationism and some blatant misconceptions followed by 'evolutionists':
http://neuroanthropology.net/2009/09/23/sympathy-for-creationists/
I'd like to add that belief in evolution is about as insane and irrational, in principle, as a belief in creation. Evolution must be understood and critically analysed at every step in order to be 'practised' correctly. It seems most people still insist on believing...
For some reason, believing in gravity sounds substantially more awkward...

Mystery Micrograph #04
Ok, I'll make it a little easier this time. Just tell me the 'type genus' of this group. And before a certain taxonomist explodes at my blasphemy there, I mean the 'famous', most prominent, genus, of this group. Which this organism may or may not be. And it's an SEM, a little easier to decipher, I hope:

(Shell of Mystery Organism. To be referenced later)
I don't think this needs a scalebar, let's say it's unicellular in a fairly normal way.
Change of rules: Since the resident protistologists (and the arachnid taxonomist who knows way too much about protists to not be suspicious...) would get this very quickly, let's bar them until Friday noon Pacific Time. This one is too easy for a free beer, but if a sane person (ie. non-protistologist/aforementioned arachnid taxonomist) gets this, you get to be famous on my blog. I know, that's just awesome! Maybe about 5 people will know you! It's almost as good as being famous in math!Or protistology itself...
Guess if no one gets it by Saturday morning I'll put up a hint...
I heard phylodeities and clade spirits like beer offerings. Just sayin' Cheers~
29.09.09: think about unicellular organisms with tests. You may have even heard of a [rather polyphyletic] group of them. Google image search could help.

(Shell of Mystery Organism. To be referenced later)
I don't think this needs a scalebar, let's say it's unicellular in a fairly normal way.
Change of rules: Since the resident protistologists (and the arachnid taxonomist who knows way too much about protists to not be suspicious...) would get this very quickly, let's bar them until Friday noon Pacific Time. This one is too easy for a free beer, but if a sane person (ie. non-protistologist/aforementioned arachnid taxonomist) gets this, you get to be famous on my blog. I know, that's just awesome! Maybe about 5 people will know you! It's almost as good as being famous in math!
Guess if no one gets it by Saturday morning I'll put up a hint...
I heard phylodeities and clade spirits like beer offerings. Just sayin' Cheers~
29.09.09: think about unicellular organisms with tests. You may have even heard of a [rather polyphyletic] group of them. Google image search could help.
Sunday Protist - Bicosoecids
So this past Mystery Micrograph has two 'half'-winners: 1. My friend's PI who can randomly pick out obscure flagellates by wandering around his lab - being unaware of this blog's (and my own) existence, we can say that's a half-win. 2. Jan, after I gave it away in the two very massive hints, so that also only counts for half. Jan is the first non-faculty member to get one of these, so congrats!
The MM answer was:
Bicosoeca

(Tong et al. 1997 Polar Biol.; Bicosoeca)
Bicosoecids are non-photosynthetic stramenopiles (see the first diagram in the Chromalveolata post); this one specifically builds itself a lorica, from which it filters the current with it's feathery anterior flagellum. (similar to choanoflagellates in some ways) They also look a bit similar to Chrysophyte Dinobryon (I spelled it right this time! =P), except that Bicosoeca lacks visible plastids. Some Bicosoecids like Pseudobodo and Cafeteria don't bother making loricas, and just anchor themselves to the substrate with one of the flagella. According to Cafeteria's Encyclopedia of Life webpage:
Despite being fairly common, these little critters are quite understudied, and very little is known about them beyond their basic morphology. Notice a common theme arising among most protist posts? Do something! We don't need THAT many people crowded on the N-terminus of some random mouse 'oncogene' (whatever that means), and there's space here!
I'll try to make next week's Mystery Micrograph a bit easier!
The MM answer was:
Bicosoeca
(Tong et al. 1997 Polar Biol.; Bicosoeca)
Bicosoecids are non-photosynthetic stramenopiles (see the first diagram in the Chromalveolata post); this one specifically builds itself a lorica, from which it filters the current with it's feathery anterior flagellum. (similar to choanoflagellates in some ways) They also look a bit similar to Chrysophyte Dinobryon (I spelled it right this time! =P), except that Bicosoeca lacks visible plastids. Some Bicosoecids like Pseudobodo and Cafeteria don't bother making loricas, and just anchor themselves to the substrate with one of the flagella. According to Cafeteria's Encyclopedia of Life webpage:
Sometimes taxonomy can be quite humourous!"As to the source of this name; it was prompted by a pink neon sign affixed to the wall of a hostelry in Roenbjerg (Denmark)which was illuminated just as the authors'were about to give up on finding a good name for one of the most significant consumers in the world."
Despite being fairly common, these little critters are quite understudied, and very little is known about them beyond their basic morphology. Notice a common theme arising among most protist posts? Do something! We don't need THAT many people crowded on the N-terminus of some random mouse 'oncogene' (whatever that means), and there's space here!
I'll try to make next week's Mystery Micrograph a bit easier!
Scientists: glorified bureaucrats?
I found this a while ago, but no one, to my knowledge, seems to have blogged about it:
Within the article is this quote:
You know how we scientists are fundamentally different from artists, how we hail from different universes and all that? I'm deeply amazed by the parallels between us! It is also nearly impossible for an artist to survive as a self-expressing independent individual - they too must subscribe to fads and funding sources (commissions, etc). They too spend most of their time freaking out over funding rather than actually practising their art. And just like our research, their art is also skewed to please thesugar daddy commissioner. We are just artists who use a different medium with different rules. But many of the common stereotypes used to ward people off foolish career choices (such as those of the fine arts) are eerily as applicable to us. After all, most of us do embark on our chosen foolish careers to satisfy some absurdly naïve inner yearnings and desires; only to find the world absolutely indifferent to our dreams.
We are all but artists-turned-businessmen.
PS: What keeps me trying to stay on this path is the realisation that I'm absolutely worthless for any other kind of job. It's been observed that having limited options often forces one to be quite happy, since there's no point in constantly reflecting upon your choice, so you adapt to it. Yay! =D
Lawrence, P. (2009). Real Lives and White Lies in the Funding of Scientific Research PLoS Biology, 7 (9) DOI: 10.1371/journal.pbio.1000197
EDIT: Speaking of academic issues, we're not so hot on the data sharing either...
Real Lives and White Lies in the Funding of Scientific ResearchGo read the article. It's scary. And seems accurate enough even to an undergrad with limited experience. (Shit, I've been jaded already before even going to grad school...)
The granting system turns young scientists into bureaucrats and then betrays them
Lawrence PA (2009) PLoS Biology 7(9): e1000197 (open access)
Within the article is this quote:
“Scientists might have had a Hippocratic oath of their own. They might have promised their gifts to mankind. But instead, I have fathered a race of inventive dwarfs who can be hired for anything.”—Bertolt Brecht “The Life of Galileo,” version by David HareTo be fair, doctors aren't exactly saints these days either, being tied up by the madness of insurance policies and a culture that deems it acceptable to SUE(!) a doctor for trying to help a patient. But at least doctors can have their own practice. You pretty much cannot set up your own business as a researcher, unless you're an engineer of sorts. Or if you blatantly contract yourself out to some organisation somewhere, which makes the research even more obviously not objective. There is hardly any real freedom or self-employment. A scientist must become a minion of their funding sources just to make ends meet.
You know how we scientists are fundamentally different from artists, how we hail from different universes and all that? I'm deeply amazed by the parallels between us! It is also nearly impossible for an artist to survive as a self-expressing independent individual - they too must subscribe to fads and funding sources (commissions, etc). They too spend most of their time freaking out over funding rather than actually practising their art. And just like our research, their art is also skewed to please the
We are all but artists-turned-businessmen.
PS: What keeps me trying to stay on this path is the realisation that I'm absolutely worthless for any other kind of job. It's been observed that having limited options often forces one to be quite happy, since there's no point in constantly reflecting upon your choice, so you adapt to it. Yay! =D
Lawrence, P. (2009). Real Lives and White Lies in the Funding of Scientific Research PLoS Biology, 7 (9) DOI: 10.1371/journal.pbio.1000197
EDIT: Speaking of academic issues, we're not so hot on the data sharing either...
Mystery Micrograph #03 HINTS
Refer to Mystery Micrograph here
1. Stramenopile
2. Non-photosynthetic
(Jan, you better get this one!)
I'll close the thing on Sunday. And it'll be really sad if so far the only MM winners are PIs. Come on guys, let's bring out the UNTENURED PRIDE!
1. Stramenopile
2. Non-photosynthetic
(Jan, you better get this one!)
I'll close the thing on Sunday. And it'll be really sad if so far the only MM winners are PIs. Come on guys, let's bring out the UNTENURED PRIDE!
I've wondered about this too...
In this week's Nature:
"Overzealous use of decimal places has wrong kind of impact"
by J. M. D. Coey 23 Sep 2009
"Overzealous use of decimal places has wrong kind of impact"
by J. M. D. Coey 23 Sep 2009
"Scientists teach students to evaluate critically the significance of their measurements, and to eschew meaningless decimal places thrown up when pocket calculators work out a quotient of two integers. So what are we to make of the recently released impact factors, including Nature's much advertised rating of '31.434' (see also http://www.nature.com/nature/about)? Has Thomson Reuters discovered a protocol that allows it to measure the impact of a journal with an accuracy of 32 p.p.m.?
Quoting this figure conveys the wrong impression — that innumerate marketing is trumping common sense at the heart of science's leading journal."
So what does a thousandth of an impact factor rating look like anyway? Also, how much does Thomson Reuters charge per point? ^_^
Mystery Micrograph #03
I'd like the genus of this one. Realising that these tend to be bloody difficult, I will allow questions - encourage them, in fact. Although I do reserve the right to not answer some...
To be referenced later; Scale - roughly ~10um across
Good luck! May the phylodeities and clade spirits be with ye!
To be referenced later; Scale - roughly ~10um across
Good luck! May the phylodeities and clade spirits be with ye!
Sunday Protist - Haptoglossa: Missile-growing spores
I totally just skipped a week, I know. But no one really noticed, right? Good! ^.^ Kind of busy with classes, setting up a seminar course, and...much neglected research. I'm beginning to forget to do treatments and such, too much stuff going on. Also, training new undergrads is a lot of work. Especially when they show up at various random times and you forget what you've explained to whom. And when they forget certain important details that you've spent a good 15min elaborating on, complete with hopefully memorable stories. Such as what happens when you hang a 2L flask on one of the plastic pegs on our drying rack, which tend to be a bit too flexible. Well, the flasks were ok, but the glass 1L cylinder - not so much... and I liked that cylinder! Oh no, I actually have 'favourite' pieces of lab equipment already...

Anyway, enough with the lab rants... remember the mystery micrograph from a good two weeks ago? The one which only one person actually bothered to guess, while the rest of you hid silently somewhere?

(Jane Robb, U of Guelph; TEM of projectile)
Admittedly, this one was kind of unfair. Turns out, it barely exists in the Encyclopedia of Life, imageless and under the very descriptive category "not assigned". And even after revealing it being an oomycete, most oomycete trees seem to utterly ignore this poor obscure organism. It's not completely obscure, as Elio at Small Things Considered beat me to posting about it a while ago, but they cover a lot of rather well-hidden life forms there. So let's welcome Haptoglossa as the guest of this week's Sunday Protist!
Before we proceed, however, let's orient ourselves phylogenetically a bit:
What are oomycetes?
Oomycetes are a non-photosynthetic fungal-like group of stramenopiles (Chromalveolata), which include Phytophthora, the potato blight 'fungus' responsible for the 19th century Irish Potato Famine, drastically impacting the course of history of both Ireland and the genetic makeup of the North American population. Oomycetes were long considered to be fungi due to the similarities in life style and morphology, thus they're still largely studied by mycologists. In fact, they were taught in the fungi section of this vastly polyphyletic course I took a couple years ago ('non-vascular plants', ha!), thereby leaving us rather confused.
One characteristic that made them stand out from their supposively fellow fungi was biflagellate zoospores - the motile spores of true fungi that produce them (ie. Chytrids) have a single posterior flagellum. Now it may seem like a trivial difference, but check this note on flagellar and basal body evolution for perspective. In short, flagellar loss (and an entirely different mechanism of flagellar propagation during cell division) is not a very common event, and can be argued to be a reasonably reliable morphological character. (Although relying on morphology alone tends to lead to complete and total chaos...) There are more fundamental differences between oomycetes and fungi, and some parallels that are quite fascinating, but I won't discuss them here.
Haptoglossa is a more or less basal oomycete, and has thus attracted attention for investigating the potential presence of relic plastid genes. (search for "Craig Bailey" in this report of the 2009 Int'l Crysophyte Symposium. Also, a whole conference full of Chrysophyte geeks - interesting thought ^.^) Here's one of the recent oomycete trees featuring Haptoglossa:

(Hakariya et al. 2007 Mycoscience; for a broader perspective, see this diagram from the Chromalveolate introduction post. This tree also shows the more 'famous' oomycetes like Phytophthora, Albugo and Peronospora, see if you can find them. Also, Cafeteria is a real genus name.)
They had three trees in that paper, but to a cell biologist they all look the same, so here's one with the highest bootstrap values. My "Phylogenetics Advisory Committee" is asleep right now. And I don't think anyone here would care much about the difference between maximum likelihood, maximum parsimony and neighbour joining analysis with respect to these particular trees. So let's move on to our guest organism.
(NB: For those interested in oomycete character evolution, there's a diagram in the appendix at the very bottom of this post. I thought it would be too distracting here...)
Haptoglossa - "Nature's ballistic missile"*
*Title of 1982 Science paper by Robb & Barron; now if only they published it in Nature instead...
You're a spore. You want to get inside a host. You really don't want to sit around hoping to get ingested or inserted by some vector. What can you do?
If you're Haptoglossa, you grow a cannon, load yourself into it, and fire away. The contraption involved looks something like this:

(Barron 1987 Mycologia; diagram of a gun cell)
Ready, set, FIRE!

(Barron 1987 Mycologia)
It seems that upon triggering, the basal vacuole expands rapidly, everting the projectile at rapid speed through the host cuticle. However, the exact mechanism of action still remains to be understood (one becomes so desensitised to this phrase in protistology...) Haptoglossa targets nematodes and rotifers, depending on the species. Once inside the host, depending on the species, some may grow into large multicellular parasites like this H.dickii:

(Hakariya et al. 2007 Mycoscience; a - spores, b - sporangium inside nematode)
To give an idea of the diversity within the genus, some species, like H.eumpens, H.heteromorpha and H.polymorpha have two distinct types of gun cells, one set of which has not been seen in action - it's exact function and behaviour remain but a mystery at the moment:

(Glockling & Beakes 2002 Fungal Genet. & Biol.; comparison of five Haptoglossa species at various life cycle stages; note the odd binucleate and 'atypical' infection cells.)
Here's the H.heteromorpha (one of the species with the mysterious alternative gun cells) life cycle in a diagram:

(Glockling & Beakes 2002 Mycologia; just follow the letters, the order is a bit counterintuitive.)
Again, it is rather puzzling why an organism would 'bother' to make two types of spores with slightly different, yet still sophisticated, firing structures. However, ridiculously off the wall complex life cycles aren't all that strange to parasites - many true fungi like 'rusts' are notorious for their particularly sadistic abuse of a mycology student's memory. Parasites can sometimes have multiple hosts if there is a seasonal cycle - ie. if you parasitise annual plants, it may actually be advantageous to be able to utilise different food sources throughout the year. However, nematodes aren't particularly seasonal, to my knowledge, so the reasons here may be something different. This could also be an ancestral feature, from the long gone days of making a living off plants, but that is rather far fetched. There may be a whole chunk of a life cycle that we are completely oblivious to at the moment...
Having quite a fetish for complex intracellular structures, I find the question of gun cell development rather fascinating. The entire structure does seem to form de novo, and a very exciting research project would be to examine the genetics and cell biology of the projectile development. While there have been observations of gun cell development, further details seem to be obscure at the moment. Here's a nice diagram based on microscopy observations:

(Glockling & Beakes 2000 Mycol Res; just sit back and savour the ability of a cell to generate seemingly limitless complexity within itself! Note that prior to this process, the spore shifts its contents into this newly generated gun cell compartment, essentially growing itself a cannon on the side.)
And another detail of the firing mechanism, and the aftermath: (the projectile on the right faces opposite of the gun cell on the left)

(Glockling & Beakes 2000 J Invert Pathol; I'm still thoroughly amazed by this mechanism!)
Now, recall that this process seems to rely on basal vacuole expansion, followed by a rapid inversion of the projectile inside out. Keeping that in mind, take a look at this sample of proximal convergence:

(source: Palaeos.org)
Microsporidia also have a clever mechanism for shooting themselves into their prey, via the posterior vacuole expanding and everting the polar tube outwards at an extremely rapid speed. There's a nice article about them on Palaeos.com, although I thought microsporidia were fairly certainly within fungi, between chytrids and zygomycetes - not sister group to Fungi. Again, my "Phylogenetics Advisory Committee: Microsporidian chapter" is unavailable at this hour...
There needs to be a protist battle game of some sort... can you imagine, firing off microsporidian and haptoglossan missles at your oponents? Only to be entangled and digested by a protoperidinian pallium? The microbial world is an endlessly fascinating place, surreal and alien to our innate perception of scale.
I've been challenged by someone to blog about Heterolobosea, which I shall do after I finish that long-overdue Neomura post. I'm not sure I can match the quality and depth of his Amoebozoa (and all the other) posts, but I'll try!
References
Barron, G. (1987). The Gun Cell of Haptoglossa mirabilis Mycologia, 79 (6) DOI: 10.2307/3807689
Hakariya, M., Hirose, D., & Tokumasu, S. (2007). A molecular phylogeny of Haptoglossa species, terrestrial peronosporomycetes (oomycetes) endoparasitic on nematodes Mycoscience, 48 (3), 169-175 DOI: 10.1007/s10267-007-0355-7
Glockling, S. (2000). An Ultrastructural Study of Sporidium Formation during Infection of a Rhabditid Nematode by Large Gun Cells of Haptoglossa heteromorpha Journal of Invertebrate Pathology, 76 (3), 208-215 DOI: 10.1006/jipa.2000.4967
Glockling, S., & Beakes, G. (2000). Video Microscopy of Spore Development in Haptoglossa heteromorpha, a New Species from Cow Dung Mycologia, 92 (4) DOI: 10.2307/3761431
Glockling, S. (2002). Ultrastructural morphogenesis of dimorphic arcuate infection (gun) cells of Haptoglossa erumpens an obligate parasite of Bunonema nematodes Fungal Genetics and Biology, 37 (3), 250-262 DOI: 10.1016/S1087-1845(02)00532-7
ROBB, E., & BARRON, G. (1982). Nature's Ballistic Missile Science, 218 (4578), 1221-1222 DOI: 10.1126/science.218.4578.1221
Appendix:
For those curious about oomycete evolution:

See Beakes & Sekimoto 2009 in Oomycete Genetics and Genomics (Eds. Lamour & Kamoun). I'm seriously annoyed by the whole arrows coming out of extant lineages/organisms thing...although "Crown Oomycetes" is rather hilarious!
Anyway, enough with the lab rants... remember the mystery micrograph from a good two weeks ago? The one which only one person actually bothered to guess, while the rest of you hid silently somewhere?

(Jane Robb, U of Guelph; TEM of projectile)
Admittedly, this one was kind of unfair. Turns out, it barely exists in the Encyclopedia of Life, imageless and under the very descriptive category "not assigned". And even after revealing it being an oomycete, most oomycete trees seem to utterly ignore this poor obscure organism. It's not completely obscure, as Elio at Small Things Considered beat me to posting about it a while ago, but they cover a lot of rather well-hidden life forms there. So let's welcome Haptoglossa as the guest of this week's Sunday Protist!
Before we proceed, however, let's orient ourselves phylogenetically a bit:
What are oomycetes?
Oomycetes are a non-photosynthetic fungal-like group of stramenopiles (Chromalveolata), which include Phytophthora, the potato blight 'fungus' responsible for the 19th century Irish Potato Famine, drastically impacting the course of history of both Ireland and the genetic makeup of the North American population. Oomycetes were long considered to be fungi due to the similarities in life style and morphology, thus they're still largely studied by mycologists. In fact, they were taught in the fungi section of this vastly polyphyletic course I took a couple years ago ('non-vascular plants', ha!), thereby leaving us rather confused.
One characteristic that made them stand out from their supposively fellow fungi was biflagellate zoospores - the motile spores of true fungi that produce them (ie. Chytrids) have a single posterior flagellum. Now it may seem like a trivial difference, but check this note on flagellar and basal body evolution for perspective. In short, flagellar loss (and an entirely different mechanism of flagellar propagation during cell division) is not a very common event, and can be argued to be a reasonably reliable morphological character. (Although relying on morphology alone tends to lead to complete and total chaos...) There are more fundamental differences between oomycetes and fungi, and some parallels that are quite fascinating, but I won't discuss them here.
Haptoglossa is a more or less basal oomycete, and has thus attracted attention for investigating the potential presence of relic plastid genes. (search for "Craig Bailey" in this report of the 2009 Int'l Crysophyte Symposium. Also, a whole conference full of Chrysophyte geeks - interesting thought ^.^) Here's one of the recent oomycete trees featuring Haptoglossa:
(Hakariya et al. 2007 Mycoscience; for a broader perspective, see this diagram from the Chromalveolate introduction post. This tree also shows the more 'famous' oomycetes like Phytophthora, Albugo and Peronospora, see if you can find them. Also, Cafeteria is a real genus name.)
They had three trees in that paper, but to a cell biologist they all look the same, so here's one with the highest bootstrap values. My "Phylogenetics Advisory Committee" is asleep right now. And I don't think anyone here would care much about the difference between maximum likelihood, maximum parsimony and neighbour joining analysis with respect to these particular trees. So let's move on to our guest organism.
(NB: For those interested in oomycete character evolution, there's a diagram in the appendix at the very bottom of this post. I thought it would be too distracting here...)
Haptoglossa - "Nature's ballistic missile"*
*Title of 1982 Science paper by Robb & Barron; now if only they published it in Nature instead...
You're a spore. You want to get inside a host. You really don't want to sit around hoping to get ingested or inserted by some vector. What can you do?
If you're Haptoglossa, you grow a cannon, load yourself into it, and fire away. The contraption involved looks something like this:

(Barron 1987 Mycologia; diagram of a gun cell)
Ready, set, FIRE!

(Barron 1987 Mycologia)
It seems that upon triggering, the basal vacuole expands rapidly, everting the projectile at rapid speed through the host cuticle. However, the exact mechanism of action still remains to be understood (one becomes so desensitised to this phrase in protistology...) Haptoglossa targets nematodes and rotifers, depending on the species. Once inside the host, depending on the species, some may grow into large multicellular parasites like this H.dickii:

(Hakariya et al. 2007 Mycoscience; a - spores, b - sporangium inside nematode)
To give an idea of the diversity within the genus, some species, like H.eumpens, H.heteromorpha and H.polymorpha have two distinct types of gun cells, one set of which has not been seen in action - it's exact function and behaviour remain but a mystery at the moment:

(Glockling & Beakes 2002 Fungal Genet. & Biol.; comparison of five Haptoglossa species at various life cycle stages; note the odd binucleate and 'atypical' infection cells.)
Here's the H.heteromorpha (one of the species with the mysterious alternative gun cells) life cycle in a diagram:
(Glockling & Beakes 2002 Mycologia; just follow the letters, the order is a bit counterintuitive.)
Again, it is rather puzzling why an organism would 'bother' to make two types of spores with slightly different, yet still sophisticated, firing structures. However, ridiculously off the wall complex life cycles aren't all that strange to parasites - many true fungi like 'rusts' are notorious for their particularly sadistic abuse of a mycology student's memory. Parasites can sometimes have multiple hosts if there is a seasonal cycle - ie. if you parasitise annual plants, it may actually be advantageous to be able to utilise different food sources throughout the year. However, nematodes aren't particularly seasonal, to my knowledge, so the reasons here may be something different. This could also be an ancestral feature, from the long gone days of making a living off plants, but that is rather far fetched. There may be a whole chunk of a life cycle that we are completely oblivious to at the moment...
Having quite a fetish for complex intracellular structures, I find the question of gun cell development rather fascinating. The entire structure does seem to form de novo, and a very exciting research project would be to examine the genetics and cell biology of the projectile development. While there have been observations of gun cell development, further details seem to be obscure at the moment. Here's a nice diagram based on microscopy observations:
(Glockling & Beakes 2000 Mycol Res; just sit back and savour the ability of a cell to generate seemingly limitless complexity within itself! Note that prior to this process, the spore shifts its contents into this newly generated gun cell compartment, essentially growing itself a cannon on the side.)
And another detail of the firing mechanism, and the aftermath: (the projectile on the right faces opposite of the gun cell on the left)
(Glockling & Beakes 2000 J Invert Pathol; I'm still thoroughly amazed by this mechanism!)
Now, recall that this process seems to rely on basal vacuole expansion, followed by a rapid inversion of the projectile inside out. Keeping that in mind, take a look at this sample of proximal convergence:

(source: Palaeos.org)
Microsporidia also have a clever mechanism for shooting themselves into their prey, via the posterior vacuole expanding and everting the polar tube outwards at an extremely rapid speed. There's a nice article about them on Palaeos.com, although I thought microsporidia were fairly certainly within fungi, between chytrids and zygomycetes - not sister group to Fungi. Again, my "Phylogenetics Advisory Committee: Microsporidian chapter" is unavailable at this hour...
There needs to be a protist battle game of some sort... can you imagine, firing off microsporidian and haptoglossan missles at your oponents? Only to be entangled and digested by a protoperidinian pallium? The microbial world is an endlessly fascinating place, surreal and alien to our innate perception of scale.
I've been challenged by someone to blog about Heterolobosea, which I shall do after I finish that long-overdue Neomura post. I'm not sure I can match the quality and depth of his Amoebozoa (and all the other) posts, but I'll try!
References
Barron, G. (1987). The Gun Cell of Haptoglossa mirabilis Mycologia, 79 (6) DOI: 10.2307/3807689
Hakariya, M., Hirose, D., & Tokumasu, S. (2007). A molecular phylogeny of Haptoglossa species, terrestrial peronosporomycetes (oomycetes) endoparasitic on nematodes Mycoscience, 48 (3), 169-175 DOI: 10.1007/s10267-007-0355-7
Glockling, S. (2000). An Ultrastructural Study of Sporidium Formation during Infection of a Rhabditid Nematode by Large Gun Cells of Haptoglossa heteromorpha Journal of Invertebrate Pathology, 76 (3), 208-215 DOI: 10.1006/jipa.2000.4967
Glockling, S., & Beakes, G. (2000). Video Microscopy of Spore Development in Haptoglossa heteromorpha, a New Species from Cow Dung Mycologia, 92 (4) DOI: 10.2307/3761431
Glockling, S. (2002). Ultrastructural morphogenesis of dimorphic arcuate infection (gun) cells of Haptoglossa erumpens an obligate parasite of Bunonema nematodes Fungal Genetics and Biology, 37 (3), 250-262 DOI: 10.1016/S1087-1845(02)00532-7
ROBB, E., & BARRON, G. (1982). Nature's Ballistic Missile Science, 218 (4578), 1221-1222 DOI: 10.1126/science.218.4578.1221
Appendix:
For those curious about oomycete evolution:
See Beakes & Sekimoto 2009 in Oomycete Genetics and Genomics (Eds. Lamour & Kamoun). I'm seriously annoyed by the whole arrows coming out of extant lineages/organisms thing...although "Crown Oomycetes" is rather hilarious!
'Crhaptophytes' and the Chromalveolate Hypothesis
Warning: This post contains copious amounts of obscure phylogeny and taxonomy. Discussed by a cell biologist. Proceed with caution.
I've probably carelessly alluded before to the Chromalveolate Hypothesis by Cavalier-Smith (eg. 2002 Curr Biol). In any case, I tend to go by the assumption it may be correct, since I'm a cell biologist and therefore required by federal law not to care about evolution. There's powerful/annoying(depending which side you're on) evidence pointing both ways, so the thing is a bit of a mess. I know, mess in protistological taxonomy? No fucking way!
Let's zoom in to one of Tom Cavalier-Smith's many warzones:

(based on Keeling et al 2005 Trends Ecol Evol;
The green dots indicate the presence of photosynthesis in respective lineages. This is just to get an idea of where these things are - members of Chromalveolata include ciliates, dinoflagellates, apicomplexans (eg. Plasmodium, responsible for malaria), diatoms, kelps, oomycetes (eg. Phytophthora, the other organism behind the Irish famine, besides H.sapiens and their sadistic and incompetent governance.) and the possibly less familiar Haptophytes (chalk in cliffs of Dover).
Now that seems like a rather diverse mix of stuff to have in one kingdom, and it is. You have multiple independent instances of multicellularity, lifestyles from parasitism to phagotrophy to photosynthesis to osmotrophy (think fungi) to mixotrophy (eg. photosynthetic predators) and beyond. It's rather hard to believe that the entire grouping may be held together by... a single red algal plastid endosymbiosis event. (TC-S 2002) And some don't. In fact, the evidence is rather strong both for and against what is called the Chromalveolate Hypothesis: where 'chromists' (stramenopiles + cryptophytes and haptophytes) and alveolates share a single secondary endosymbiosis event.
If the Chromalv. hypothesis is accurate, you would expect many lineages to be photosynthetic or contain relic plastids. Furthermore, you'd expect lineages devoid of plastids to at least contain some relic plastid-derived genes in their genome. Those characters should also point towards a single origin, as opposed to two or more independent endosymbiosis events (eg. from different red algae).
I've prepared an overview of what the Chromalv. hypothesis 'looks like', hopefully not plagued by too many inaccuracies:

(This almost looks like a TC-S diagram. I guess that's just inevitable. Red - groups containing photosynthetic lineages with a red-algal derived plastid; Green - group with green algal secondarily derived plastid. EDIT: And that's KAtablepharids, not Ketablepharids...EDIT#2: Forgot the excavates - place them somewhere between the archaeplastid-chromalv node and the root...)
Going clade by clade, some evidence that supports single chromalv. plastid origin is:
- Apicomplexa, a vast group of intracellular parasites such as not-so-friendly(to us) critters like Plasmodium and Toxoplasma, have been found to posess reduced plastids, called apicoplasts (eg. reviewed in Waller & McFadden 2005 Curr Issues Mol Biol). Malaria turns out to be an algal disease. There's plenty of other examples of algae-turned-parasites, but we've got a TC-S hypothesis to cover...
- Basal to Apicomplexa is a photosynthetic alga called Chromera, with a red-algal-derived plastid, which further supports an algal origin of 'Apies'. (Moore, Oborník, Janouškovec et al. 2008 Nature)
- Dinoflagellates, Ochrophytes (group containing kelps and diatoms), Cryptomonads and Haptophytes all have photosynthetic members with a certain red-algal derived plastid. Now, the fuss is about whether they all got their plastids once, with the plastid-less lineages having lost them through time, or multiple times within the Chromalveolate kingdom.
- (more evidence is discussed in Keeling 2009 JEM)
Refer to the diagram below. Endosymbiosis is accepted by everyone but Marguilis to be a very unlikely event, and therefore very unparsimonious to postulate for every photosynthetic lineage you see. You would expect some lineages to lose their photosynthetic ability, and even their plastids altogether. However, since endosymbiosis usually results in gene transfer to the host, you should be able to find plastid-derived genes in most lineages. This means that both plastid-bearing and plastid-less lineages should be distributed fairly haphazardly, without too much non-photosynthetic stuff clumping around basally. Unfortunately, that is annoyingly not the case entirely:
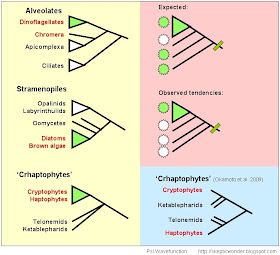
Prior to the discovery of Chromera and apicoplasts, the Chromalv. hypothesis was rather weak in Alveolata, with ciliates and apies both being non-photosynthetic. Furthermore, the basal lineages of Stramenopiles are also non-photosynthetic, with things like 'fungal' oomycetes, labyrinthulids, opalinids ('ciliated'-looking frog gut endosymbionts), and parasites and gut commensals like Blastocystis and Proteromonas that seldom see the light of day. Among the Crypto-Hapto clade, it also seemed like the non-photosynthetic stuff like Katablepharids and Telonemids were basal to a monophyletic crypto-hapto grouping. When there's a long stretch of basal lineages devoid of a certain trait, it becomes more unlikely for that trait to have been acquired early on. Compare the bottom two trees - which topology is more likely to agree with an ancient common plastid origin?
It is fairly certain that the alveolate clade shares a common endosymbiotic event - there are even some seemingly-algal derived genes in ciliates, although that remains to be confirmed (Prieto et al. 2008 Curr Biol). Among the Stramenopiles, things seem to be clearing up a bit - some Oomycetes seem to have putative plastid-derived genes, for example. And something nice was recently found in the Cryptomonad-Haptophyte group.
Cryptomonads and haptophytes are the more familiar members (well, relatively) of the newly established Hacrobia, or 'Crhaptophytes'. Cryptomonads are fairly ubiquitous little algae with a characteristic gullet lined with ejectosomes - organelles that basically sting their prey. Cryptomonads are special in that they have nucleomorphs - highly reduced nuclei left behind from the red algal secondary endosymbiosis. The crypto plastids actually have a remnant red algal nucleus in the remainder of the endosymbionts cytoplasm. Nucleomorphs are known in one other organism - chlorarachniophytes, which have a green algal secondary endosymbiont (see the big chromalveolata diagram above). Due to the size constraints and reduced need for making own proteins (since the host can provide them), the nucleomorph genomes have some rather interesting features, which should be covered at a later date.
Haptophytes are generally spherical things, characterised by having a protrustion (haptonema) between its flagella, which it may use to catch prey. The more famous haptos are covered in little scales (coccoliths), and are great contributors to sedimentation, often resulting in...chalk! Coccolith-bearing haptos are extremely tiny organisms, which can be seen from space. A taxonomic mess resulted from there being seemingly two types of haptos: heterococcoliths and holococcoliths. They were long considered asexual until Geisen et al. 2002 reported a half-hetero- half homococcolith-bearing haptophyte, which led to the discovery of a sexual lifecycle involving a diploid heterococcolith and a haploid homococcolith stage, illustrated here. It's very interesting how traditionally 'asexual' lineages seem to be foung otherwise, one by one...


(Source: Micro*scope (crypto); http://www.geo.uni-bremen.de/cocco/(hapto))
You may be wondering where the 'crhaptophytes' come from. Time for a piece of lab folklore told to me by a postdoc:
Once upon a time, when the support for the Cryptophyte-Haptophyte clade grew to near certainty, one of the authors of that paper coined the term 'Crhaptophytes', because it's a rather apt description of the grouping, and sounds pretty awesome. And they are tiny hyperactive pieces of crap. Apparently 'Crhaptophyte' didn't fly well with the ICZN/ICBN taxonazis, since their true intention is to make biological nomenclature living hell to deal with, and impossible to stay awake through. In short, a sleeping hell. So they had to come up with another name, and Hacrobia (ewww) it was.Of course, postdocs may not be the most reliable people on the planet when it comes to storytelling to potentially gullible undergrads, and academic gossip in general is very oddly like any other kind of gossip. However, given the cast of characters involved, it's not altogether impossible.
Since I really prefer the term Crhaptophyte, I'll encourage its use wherever possible. You know, taxonomy should be fun! Wow, 'taxonomy' and 'fun' in the same sentence, without a negation.What has the world come to...!
Anyway, back to our story: Lets meet Roombia, named after a...robotic vacuum cleaner! (Okamoto et al. 2009 PLoS ONE)

(Okamoto et al. 2009 PLoS ONE - Roombia truncata; scalebar - 5um)
Sometimes a new organism is found that straightens out and stabilises messy trees (or utterly wrecks well-accepted ones...) The monophyly of our 'Chraptophytes' wasn't completely established until fairly recently, and the relationships of the organisms within that group were also murky. Sometimes the sequences just don't like you and spew out trees that make no sense.

If you go back up to this diagram, the relevance of this finding to the Chromalv. hypothesis is evident in the bottom two trees: the placement of some non-photosynthetic lineages among both of two distinct clades - one with haptos and one with cryptos - takes care of one more troublesome clade with long stretches of basal non-photosynthetic crap chomping away at the likelihood of a single origin. However, there's still the issue of whether Hacrobia is sister to Stramenopiles + Alveolates (good for Chromalv.) or Archaeplastids (bad); as well as establishing whether the "stramalveolate" (can we call it that for now?) plastids and Hacrobia plastids are likely to share a common origin between them.
So it looks like the chromalveolate hypothesis is doing a little bit better, now that not all the basal lineages are non-photosynthetic; multiple plastid loss seems more plausible. However, if you look at one of the broader trees in the paper, it seems to show a pretty well-supported (bootstrap of 0.98) branching of Crhaptos with Archaeplastida. That would be very bad (eg. . However, this is only one gene, Hsp90, and the paper doesn't really discuss that anywhere. Furthermore, my sources inform me that the root(Archaeplastids, Crhapto, (Rhiz, Stramenopiles, Alveolates)) branching seems more likely at the moment. In fact, the little barely-noticeable hyphens under the branchings mean that less that half of the trees support that topology, so ignore everything I just said about archaeplastids and crhaptos. This is to remind us that you can't infer anything from looking at a single tree - biological data is inherently messy, no matter how pretty and crisp and clean the diagram...
By the way, this is by far not all there is to work around the Chromalveolate Hypothesis. There's so much more data pointing in both directions, but I have neither the time nor the qualification to write about it all... and hopefully I haven't butchered too much stuff in this post.
I guess the main thing to take home from all this is how messy and complicated the task of establishing relationships between organisms can be. One must be very skeptical of any models based on single evidence, such as single gene trees or morphology. Or even multiple gene trees, if other data is ignored. Weird shit happens in evolution, and some mysterious event in the past can seriously screw up our perception of the present. As can yet-to-be-discovered characters and phenomena: the Archaezoa were doing quite well until most of the 'amitochondriate' lineages were found to actually contain relics of mitochondrial presence. New data can completely turn trees upside down, roots left dangling all over the place. New data can also strengthen a model. You just never really know, and have to rely on logic, likelihoods and, admittedly, a fair amount of hunches. One must be flexible in the face of new evidence, yet skeptical of the interpretations and validity thereof. Such is life in the fragile wedlock of skepticism and wonder that is science.
PS: Image searches can lead to strange places. Like CreationWiki. Owww. I can feel parts of my brain rotting away and draining out of various cranial orifices:
"All haptophytes are asexual" WRONG. And WHAT THE FLYING FUCK does Blastocystis have to do with Apicomplexa?! Oh right, we reject evolution, therefore phylogeny no longer matters, you polyphylophilic* ignorant morons. Note the absense of references: everything seems to be stolen from Wikipedia et al. Shocking. AND OH MY FSM: "Baraminology"! o_O I'll just leave it at that...
*Adj. Having a fetish for knowingly using polyphyletic lineages. A known side effect of phylogenetic ignorance. Actually, I feel kinda bad bullying the 'epistemologically challenged'. It's more fun to pick on bad PNAS papers instead...
PPS: I'm a nerd: [after subscribing to Trends Ecol Evol feed via Reader]"I've singlehandedly just raised TrEE's IF by a good 10 points! "
My friend (hi!=D) is just evil: "send there a paper to pull it back."
[followed by more impact factor jokes...]
EDIT: David and Jan, thanks for the corrections!
Cavalier-Smith, T. (2002). Chloroplast Evolution: Secondary Symbiogenesis and Multiple Losses Current Biology, 12 (2) DOI: 10.1016/S0960-9822(01)00675-3
KEELING, P., BURGER, G., DURNFORD, D., LANG, B., LEE, R., PEARLMAN, R., ROGER, A., & GRAY, M. (2005). The tree of eukaryotes Trends in Ecology & Evolution, 20 (12), 670-676 DOI: 10.1016/j.tree.2005.09.005
KEELING, P. (2009). Chromalveolates and the Evolution of Plastids by Secondary Endosymbiosis Journal of Eukaryotic Microbiology, 56 (1), 1-8 DOI: 10.1111/j.1550-7408.2008.00371.x
Moore, R., Oborník, M., Janouškovec, J., Chrudimský, T., Vancová, M., Green, D., Wright, S., Davies, N., Bolch, C., Heimann, K., Šlapeta, J., Hoegh-Guldberg, O., Logsdon, J., & Carter, D. (2008). A photosynthetic alveolate closely related to apicomplexan parasites Nature, 451 (7181), 959-963 DOI: 10.1038/nature06635
Okamoto, N., Chantangsi, C., Horák, A., Leander, B., & Keeling, P. (2009). Molecular Phylogeny and Description of the Novel Katablepharid Roombia truncata gen. et sp. nov., and Establishment of the Hacrobia Taxon nov PLoS ONE, 4 (9) DOI: 10.1371/journal.pone.0007080
REYESPRIETO, A., MOUSTAFA, A., & BHATTACHARYA, D. (2008). Multiple Genes of Apparent Algal Origin Suggest Ciliates May Once Have Been Photosynthetic Current Biology, 18 (13), 956-962 DOI: 10.1016/j.cub.2008.05.042
The joys of old literature...
Call me immature, but I succumbed to a 5min laughing fit upon seeing this while digging through 17-18th century articles in JSTOR*:

(van Leeuwenhoek 1711 Phil. Transac. 27:316-320)
Also, I can't help but mentally read this stuff with a lisp, thanks to the f-shaped s's...
But "they call it afsucking ram, because it had fsuck'd it's Dam a little before it was killed" is just insane...
*That is what normal people do at 12.30am for fun, right?
(van Leeuwenhoek 1711 Phil. Transac. 27:316-320)
Also, I can't help but mentally read this stuff with a lisp, thanks to the f-shaped s's...
But "they call it a
*That is what normal people do at 12.30am for fun, right?
A new low: anime "Erythrops"
Oh my FSM, what have I created?

Erythrops the [cyclopsoid] Erythropsidinium. Now I gotta dig up that Greuet 1977 paper again to see if this looks a bit less awkward in mitosis... I mean, uhhh, focus on my classes and study my biochem notes. Yeah. That.
Hey, when they realise their mistake and kick me out of academia, I can totally become a protist comic artist! Now that's a job in high demand out in the real world...
As for Sunday Protist, I'm waiting for a certain someone to reappear online and remedy his guess. We're a bit low on Dalhousie traffic lately... I'm looking at you, Ancyromonas geek!
Anyway, class in 7.5h... totally gonna attend it! Nothing! can be more fun and exciting! than 8am! biochem! Wheeee! Yay!
Erythrops the [cyclopsoid] Erythropsidinium. Now I gotta dig up that Greuet 1977 paper again to see if this looks a bit less awkward in mitosis... I mean, uhhh, focus on my classes and study my biochem notes. Yeah. That.
Hey, when they realise their mistake and kick me out of academia, I can totally become a protist comic artist! Now that's a job in high demand out in the real world...
As for Sunday Protist, I'm waiting for a certain someone to reappear online and remedy his guess. We're a bit low on Dalhousie traffic lately... I'm looking at you, Ancyromonas geek!
Anyway, class in 7.5h... totally gonna attend it! Nothing! can be more fun and exciting! than 8am! biochem! Wheeee! Yay!
Mystery Micrograph #02 - last call!
Just guess something!
Should be enough hints here:
Hint #1: predation
Hint #2: protist
Scale: thing is ~5-10um long
Hint #3: parasitism
Hint #4: I have a fetish for complex cell structure
Hint #5: multicellular forms exist in this group
Hint #6: chromalveolate
Last hint: hyphal stramenopile (this should be easy-ish now!)
Genus or cell type name would do!
Extension until tomorrow evening, where I aim to post the answer, and the Sunday Protist...
Should be enough hints here:
Hint #1: predation
Hint #2: protist
Scale: thing is ~5-10um long
Hint #3: parasitism
Hint #4: I have a fetish for complex cell structure
Hint #5: multicellular forms exist in this group
Hint #6: chromalveolate
Last hint: hyphal stramenopile (this should be easy-ish now!)
Genus or cell type name would do!
Extension until tomorrow evening, where I aim to post the answer, and the Sunday Protist...
100th post!
100th post, yay! Time for something silly, to celebrate:
Let's meet our new mascots, Cer Ratium (pun avoidance order in effect) and [insert witty name here] the Protoperidinium:


(ARGH my shitty computer just can't handle the elite memory whore that is ImageJ...)
Cer Ratium has a special attack: for the anime geeks among us, remember Lust from Fullmetal Alchemist? Yeah... this thing is gonna grow unrealistically long finger[nail] things to pierce opponents with...
And Protoperidinium ("Protoper"?) has this epic pallium attack where it casts a pseudopodious veil upon its opponents and digests them. Size advantages are useless against this one!
And now some of the other cast members: (mostly borne of lecture note margin doodles...)

Left -> right: T-phage riding E.coli, Psi, perpetually hammered EtOH, Partial [derivative], hNu, perpetually abused Neuron, a couple dimensions (R3 with the Projection Lance, mwahaha), Caffeine, cute lil' Kinesin and Dynein chillin on a 'tube (microtubule)... Dynein is holding up a clathrin-coated vesicle.
("Эх, прокачу!" is a reference to a Russian novel, Золотой Телёнок...)
Note: I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd...
Let's meet our new mascots, Cer Ratium (pun avoidance order in effect) and [insert witty name here] the Protoperidinium:
(ARGH my shitty computer just can't handle the elite memory whore that is ImageJ...)
Cer Ratium has a special attack: for the anime geeks among us, remember Lust from Fullmetal Alchemist? Yeah... this thing is gonna grow unrealistically long finger[nail] things to pierce opponents with...
And Protoperidinium ("Protoper"?) has this epic pallium attack where it casts a pseudopodious veil upon its opponents and digests them. Size advantages are useless against this one!
And now some of the other cast members: (mostly borne of lecture note margin doodles...)
Left -> right: T-phage riding E.coli, Psi, perpetually hammered EtOH, Partial [derivative], hNu, perpetually abused Neuron, a couple dimensions (R3 with the Projection Lance, mwahaha), Caffeine, cute lil' Kinesin and Dynein chillin on a 'tube (microtubule)... Dynein is holding up a clathrin-coated vesicle.
("Эх, прокачу!" is a reference to a Russian novel, Золотой Телёнок...)
Note: I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd. I am not a nerd...
A quick note on flagella, and their evolution
First off, 'flagella' and 'cilia' tend to be used interchangeably. I prefer to call them flagella, out of habit, but there's some who argue 'flagellum' should be reserved for bacteria, who have a fundamentally different system from us; while we have 'cilia'. Another note: 'flagella' is spelled with two l's, 'cilia' with one. Took me about two months of protistology to learn that. (also, I consistently spelled 'axopodia' as 'auxopodia', thanks to a plant biology research background. Curse you, auxin!)
Interestingly the flagellar structures seem to be fairly conserved in evolution, and are often used in taxonomy. Most eukaryotes are fundamentally biflagellate, meaning their flagellar systems, whatever they are, are likely derived from modifications of an ancestral biflagellate form, and retaining the double basal bodies. Flagella can be lost, but the basal bodies that anchor them tend to remain behind. Conversely, basal bodies can be duplicated, as they have, for example, in parabasalia, which are tetraflagellate; entire basal body units (kinetids) can also be multiplied, up to extremes such as in ciliates. (the developmental organisation of ciliate flagella is an endlessly fascinating subject, and if all goes well, would be my research focus after BSc. =D *knocks onher head wood)
In contrast, a few eukaryotes have what is fundamentally a single flagellum - those are unikonts, which include amoebozoa (eg. cellular slime moulds) and opisthokonts (ass-tails, eg. fungi, choanoflagellates...and us). It is intuitive to think of flagella as propelling the organism forward. But not everything is about sperm: most eukaryotic organisms actually pull themselves by flagellar motion, thereby defining the location of the flagellum as the anterior end, rather than posterior. Another distinction is between isokonts (equal flagella) and heterokonts (unequal flagella) - in the former, the two flagella are structurally identical, whereas in the latter they differ, often with little protrusions (mastigonemes) lining one of them.
Actually, scratch everything I just said about opisthokonts and amoebozoa being unikonts together. Missed a memo... there's this amitochondriate amoeba Breviata (TCoO post here and picture here), previously of uncertain placement or classified as an archamoeba. Despite having a single flagellum, it seems to have a double basal body, one of them unflagellated (Walker et al. 2006 JEM). Turns out that evidence suggests it's a basal amoebozoan, which would kill TC-S' unikont/bikont division (indicated in grey below):

(Roger & Simpson 2009 Curr Biol; numbers indicate ancestral number of basal bodies/flagellar unit, asterisk indicates one basal body is unflagellated, and the 2+ in Breviata indicates there may have been more that two basal bodies/unit.)
So to summarise:
kinetid - unit of basal bodies + flagella; not all basal bodies must have a flagellum (but the flagella must be anchored to a basal body each)
opisthokont - organisms with posterior flagellation; most eukaryotes have flagella at the front of their movement.
heterokont - both (or more) flagella structurally different
isokont - both (or more) flagella are the same
unikont - organisms with single basal body/flagellum per kinetid
bikont - organisms with double (or more) basal body per kinetid
mastigonemes - little protrusions regularly lining a flagellum; for increasing a flagellum's surface area.
centriole/basal body - generally interchangeable
cilium/flagellum - generally interchangeable
(kont means tail, by the way)
I noticed I throw those terms a lot in other posts without really explaining them; so hopefully this post can be some sort of reference, just in case!
There's more to it, but someone has some protist-oriented microscopy for me to do. I love Saturday nights!
Roger, A., & Simpson, A. (2009). Evolution: Revisiting the Root of the Eukaryote Tree Current Biology, 19 (4) DOI: 10.1016/j.cub.2008.12.032
WALKER, G., DACKS, J., & MARTIN EMBLEY, T. (2006). Ultrastructural Description of Breviata anathema, N. Gen., N. Sp., the Organism Previously Studied as "Mastigamoeba invertens" The Journal of Eukaryotic Microbiology, 53 (2), 65-78 DOI: 10.1111/j.1550-7408.2005.00087.x
Interestingly the flagellar structures seem to be fairly conserved in evolution, and are often used in taxonomy. Most eukaryotes are fundamentally biflagellate, meaning their flagellar systems, whatever they are, are likely derived from modifications of an ancestral biflagellate form, and retaining the double basal bodies. Flagella can be lost, but the basal bodies that anchor them tend to remain behind. Conversely, basal bodies can be duplicated, as they have, for example, in parabasalia, which are tetraflagellate; entire basal body units (kinetids) can also be multiplied, up to extremes such as in ciliates. (the developmental organisation of ciliate flagella is an endlessly fascinating subject, and if all goes well, would be my research focus after BSc. =D *knocks on
In contrast, a few eukaryotes have what is fundamentally a single flagellum - those are unikonts, which include amoebozoa (eg. cellular slime moulds) and opisthokonts (ass-tails, eg. fungi, choanoflagellates...and us). It is intuitive to think of flagella as propelling the organism forward. But not everything is about sperm: most eukaryotic organisms actually pull themselves by flagellar motion, thereby defining the location of the flagellum as the anterior end, rather than posterior. Another distinction is between isokonts (equal flagella) and heterokonts (unequal flagella) - in the former, the two flagella are structurally identical, whereas in the latter they differ, often with little protrusions (mastigonemes) lining one of them.
Actually, scratch everything I just said about opisthokonts and amoebozoa being unikonts together. Missed a memo... there's this amitochondriate amoeba Breviata (TCoO post here and picture here), previously of uncertain placement or classified as an archamoeba. Despite having a single flagellum, it seems to have a double basal body, one of them unflagellated (Walker et al. 2006 JEM). Turns out that evidence suggests it's a basal amoebozoan, which would kill TC-S' unikont/bikont division (indicated in grey below):

(Roger & Simpson 2009 Curr Biol; numbers indicate ancestral number of basal bodies/flagellar unit, asterisk indicates one basal body is unflagellated, and the 2+ in Breviata indicates there may have been more that two basal bodies/unit.)
So to summarise:
kinetid - unit of basal bodies + flagella; not all basal bodies must have a flagellum (but the flagella must be anchored to a basal body each)
opisthokont - organisms with posterior flagellation; most eukaryotes have flagella at the front of their movement.
heterokont - both (or more) flagella structurally different
isokont - both (or more) flagella are the same
unikont - organisms with single basal body/flagellum per kinetid
bikont - organisms with double (or more) basal body per kinetid
mastigonemes - little protrusions regularly lining a flagellum; for increasing a flagellum's surface area.
centriole/basal body - generally interchangeable
cilium/flagellum - generally interchangeable
(kont means tail, by the way)
I noticed I throw those terms a lot in other posts without really explaining them; so hopefully this post can be some sort of reference, just in case!
There's more to it, but someone has some protist-oriented microscopy for me to do. I love Saturday nights!
Roger, A., & Simpson, A. (2009). Evolution: Revisiting the Root of the Eukaryote Tree Current Biology, 19 (4) DOI: 10.1016/j.cub.2008.12.032
WALKER, G., DACKS, J., & MARTIN EMBLEY, T. (2006). Ultrastructural Description of Breviata anathema, N. Gen., N. Sp., the Organism Previously Studied as "Mastigamoeba invertens" The Journal of Eukaryotic Microbiology, 53 (2), 65-78 DOI: 10.1111/j.1550-7408.2005.00087.x
Which opisthokont amoeboflagellate is this!?
While looking through the developmental biol lecture notes yesterday, I came across this image:

(Developmental Biology 8th ed. Gilbert fig 7.2)
So I sit there, staring at it, thinking: "Ok, so it's an amoeboflagellate opisthokont capable of transforming between the two. Shit, what kind of protist is this!? Not a Heterolobosean (those famously form flagella+centrioles de novo) due to its posterior flagellum and continuously present centrioles. Can't be a nucleariid or ichthyosporid or anything..." for a good 5min, getting frustrating that I couldn't recognise this protist... (also wondering why there were protists in a classical developmental biol class)
...and then I read 'sperm head'.
I'm so gonna fail this class in 3...2...1...
Well, actually I was right: this -is- an amoeboflagellate opisthokont with de novo flagellar formation. To a cladist, we'd be protists too!

(Developmental Biology 8th ed. Gilbert fig 7.2)
So I sit there, staring at it, thinking: "Ok, so it's an amoeboflagellate opisthokont capable of transforming between the two. Shit, what kind of protist is this!? Not a Heterolobosean (those famously form flagella+centrioles de novo) due to its posterior flagellum and continuously present centrioles. Can't be a nucleariid or ichthyosporid or anything..." for a good 5min, getting frustrating that I couldn't recognise this protist... (also wondering why there were protists in a classical developmental biol class)
...and then I read 'sperm head'.
I'm so gonna fail this class in 3...2...1...
Well, actually I was right: this -is- an amoeboflagellate opisthokont with de novo flagellar formation. To a cladist, we'd be protists too!
Finger-growing dinoflagellate (cute!)
Anyway, apparently Ceratium ranipes, a photosynthetic dinoflagellate, decided to grow plastid-stuffed 'fingers' during daylight:

(Pizay et al. 2009 Protist, in press; light period)
And retracts them back in for the night:

(Pizay et al. 2009 Protist, in press; dark period)
In case you're not convinced these are the same organism:

(Pizay et al. 2009 Protist, in press; sequence from a single individual removed from light, T measures minutes of darkness) (a video of that would be so awesome...!)
Chloroplasts fluoresce red when hit by UV light (see my own example with a diatom); take a look at those fingers:


(Pizay et al. 2009 Protist, in press. Left: formalin-preserved C.ranipes with a daytime morphology; the inset shows UV autofluorescense of the plastids: note their concentration in the 'fingers'. The bluish/whitish subinset shows Calcofluor White staining, which indicates the presence of thecal plates on the fingers. Right: transitional morphology at the end of the day: note how the plastids migrated inwards away from the fingers.)
This raises some cell biology-related questions: how is plastid movement coordinated and regulated? What does the genetic developmental pathway look like for those fingers, and how does it interact with whatever immediately respond to light? More importantly, why does this thing seemingly waste its time growing and retracting fingers, when it could have just kept them protruded during dark hours?
Could be just a low cost glitch in the system, or perhaps there is something to it. After all, perhaps it wouldn't take much to lose the finger retraction ability - so is there some cost when that happens, thereby keeping this process going? Sinking may have something to do with it - many planktonic algae sink for the night and float back up during the day. Fingers may drastically slow down the sinking speed. However, there's no data yet showing any vertical migration in C.ranipes (Pizay et al. 2009 Protist). Could be a relic from an ancestor that did sink, but then why hasn't this behaviour been found earlier, and in more dinos?
Another idea in the same paper is that the fingers get in the way of directed swimming; during daylight hours, you sacrifice your swimming ability for a larger photosynthetic capacity, but it may be advantageous to put away the tackle in the absense of light.
This reminds me of two things: 1) plant leaves - increasing the surface area exposed to surroundings for gas exchange, as well as the area exposed to light. Sort of a convergence. 2) Many 'radiolaria' have algal symbionts they use for photosynthesis, and they too spread them out towards the tips of the host's filopodia during daytime, and retract them inwards for the night. So whatever you do, don't dangle your plastids in plain view when they're not in use.
---
Pizay, M., Lemée, R., Simon, N., Cras, A., Laugier, J., & Dolan, J. (2009). Night and Day Morphologies in a Planktonic Dinoflagellate Protist DOI: 10.1016/j.protis.2009.04.003