
Warning: This post contains copious amounts of obscure phylogeny and taxonomy. Discussed by a cell biologist. Proceed with caution.
I've probably carelessly alluded before to the Chromalveolate Hypothesis by Cavalier-Smith (eg. 2002 Curr Biol). In any case, I tend to go by the assumption it may be correct, since I'm a cell biologist and therefore required by federal law not to care about evolution. There's powerful/annoying(depending which side you're on) evidence pointing both ways, so the thing is a bit of a mess. I know, mess in protistological taxonomy? No fucking way!
Let's zoom in to one of Tom Cavalier-Smith's many warzones:

(based on Keeling et al 2005 Trends Ecol Evol;
The green dots indicate the presence of photosynthesis in respective lineages. This is just to get an idea of where these things are - members of Chromalveolata include ciliates, dinoflagellates, apicomplexans (eg. Plasmodium, responsible for malaria), diatoms, kelps, oomycetes (eg. Phytophthora, the other organism behind the Irish famine, besides H.sapiens and their sadistic and incompetent governance.) and the possibly less familiar Haptophytes (chalk in cliffs of Dover).
Now that seems like a rather diverse mix of stuff to have in one kingdom, and it is. You have multiple independent instances of multicellularity, lifestyles from parasitism to phagotrophy to photosynthesis to osmotrophy (think fungi) to mixotrophy (eg. photosynthetic predators) and beyond. It's rather hard to believe that the entire grouping may be held together by... a single red algal plastid endosymbiosis event. (TC-S 2002) And some don't. In fact, the evidence is rather strong both for and against what is called the Chromalveolate Hypothesis: where 'chromists' (stramenopiles + cryptophytes and haptophytes) and alveolates share a single secondary endosymbiosis event.
If the Chromalv. hypothesis is accurate, you would expect many lineages to be photosynthetic or contain relic plastids. Furthermore, you'd expect lineages devoid of plastids to at least contain some relic plastid-derived genes in their genome. Those characters should also point towards a single origin, as opposed to two or more independent endosymbiosis events (eg. from different red algae).
I've prepared an overview of what the Chromalv. hypothesis 'looks like', hopefully not plagued by too many inaccuracies:

(This almost looks like a TC-S diagram. I guess that's just inevitable. Red - groups containing photosynthetic lineages with a red-algal derived plastid; Green - group with green algal secondarily derived plastid. EDIT: And that's KAtablepharids, not Ketablepharids...EDIT#2: Forgot the excavates - place them somewhere between the archaeplastid-chromalv node and the root...)
Going clade by clade, some evidence that supports single chromalv. plastid origin is:
- Apicomplexa, a vast group of intracellular parasites such as not-so-friendly(to us) critters like Plasmodium and Toxoplasma, have been found to posess reduced plastids, called apicoplasts (eg. reviewed in Waller & McFadden 2005 Curr Issues Mol Biol). Malaria turns out to be an algal disease. There's plenty of other examples of algae-turned-parasites, but we've got a TC-S hypothesis to cover...
- Basal to Apicomplexa is a photosynthetic alga called Chromera, with a red-algal-derived plastid, which further supports an algal origin of 'Apies'. (Moore, Oborník, Janouškovec et al. 2008 Nature)
- Dinoflagellates, Ochrophytes (group containing kelps and diatoms), Cryptomonads and Haptophytes all have photosynthetic members with a certain red-algal derived plastid. Now, the fuss is about whether they all got their plastids once, with the plastid-less lineages having lost them through time, or multiple times within the Chromalveolate kingdom.
- (more evidence is discussed in Keeling 2009 JEM)
Refer to the diagram below. Endosymbiosis is accepted by everyone but Marguilis to be a very unlikely event, and therefore very unparsimonious to postulate for every photosynthetic lineage you see. You would expect some lineages to lose their photosynthetic ability, and even their plastids altogether. However, since endosymbiosis usually results in gene transfer to the host, you should be able to find plastid-derived genes in most lineages. This means that both plastid-bearing and plastid-less lineages should be distributed fairly haphazardly, without too much non-photosynthetic stuff clumping around basally. Unfortunately, that is annoyingly not the case entirely:
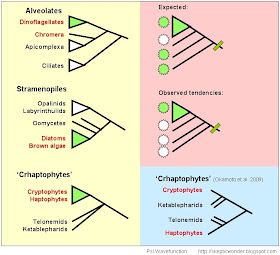
Prior to the discovery of Chromera and apicoplasts, the Chromalv. hypothesis was rather weak in Alveolata, with ciliates and apies both being non-photosynthetic. Furthermore, the basal lineages of Stramenopiles are also non-photosynthetic, with things like 'fungal' oomycetes, labyrinthulids, opalinids ('ciliated'-looking frog gut endosymbionts), and parasites and gut commensals like Blastocystis and Proteromonas that seldom see the light of day. Among the Crypto-Hapto clade, it also seemed like the non-photosynthetic stuff like Katablepharids and Telonemids were basal to a monophyletic crypto-hapto grouping. When there's a long stretch of basal lineages devoid of a certain trait, it becomes more unlikely for that trait to have been acquired early on. Compare the bottom two trees - which topology is more likely to agree with an ancient common plastid origin?
It is fairly certain that the alveolate clade shares a common endosymbiotic event - there are even some seemingly-algal derived genes in ciliates, although that remains to be confirmed (Prieto et al. 2008 Curr Biol). Among the Stramenopiles, things seem to be clearing up a bit - some Oomycetes seem to have putative plastid-derived genes, for example. And something nice was recently found in the Cryptomonad-Haptophyte group.
Cryptomonads and haptophytes are the more familiar members (well, relatively) of the newly established Hacrobia, or 'Crhaptophytes'. Cryptomonads are fairly ubiquitous little algae with a characteristic gullet lined with ejectosomes - organelles that basically sting their prey. Cryptomonads are special in that they have nucleomorphs - highly reduced nuclei left behind from the red algal secondary endosymbiosis. The crypto plastids actually have a remnant red algal nucleus in the remainder of the endosymbionts cytoplasm. Nucleomorphs are known in one other organism - chlorarachniophytes, which have a green algal secondary endosymbiont (see the big chromalveolata diagram above). Due to the size constraints and reduced need for making own proteins (since the host can provide them), the nucleomorph genomes have some rather interesting features, which should be covered at a later date.
Haptophytes are generally spherical things, characterised by having a protrustion (haptonema) between its flagella, which it may use to catch prey. The more famous haptos are covered in little scales (coccoliths), and are great contributors to sedimentation, often resulting in...chalk! Coccolith-bearing haptos are extremely tiny organisms, which can be seen from space. A taxonomic mess resulted from there being seemingly two types of haptos: heterococcoliths and holococcoliths. They were long considered asexual until Geisen et al. 2002 reported a half-hetero- half homococcolith-bearing haptophyte, which led to the discovery of a sexual lifecycle involving a diploid heterococcolith and a haploid homococcolith stage, illustrated here. It's very interesting how traditionally 'asexual' lineages seem to be foung otherwise, one by one...


(Source: Micro*scope (crypto); http://www.geo.uni-bremen.de/cocco/(hapto))
You may be wondering where the 'crhaptophytes' come from. Time for a piece of lab folklore told to me by a postdoc:
Once upon a time, when the support for the Cryptophyte-Haptophyte clade grew to near certainty, one of the authors of that paper coined the term 'Crhaptophytes', because it's a rather apt description of the grouping, and sounds pretty awesome. And they are tiny hyperactive pieces of crap. Apparently 'Crhaptophyte' didn't fly well with the ICZN/ICBN taxonazis, since their true intention is to make biological nomenclature living hell to deal with, and impossible to stay awake through. In short, a sleeping hell. So they had to come up with another name, and Hacrobia (ewww) it was.Of course, postdocs may not be the most reliable people on the planet when it comes to storytelling to potentially gullible undergrads, and academic gossip in general is very oddly like any other kind of gossip. However, given the cast of characters involved, it's not altogether impossible.
Since I really prefer the term Crhaptophyte, I'll encourage its use wherever possible. You know, taxonomy should be fun! Wow, 'taxonomy' and 'fun' in the same sentence, without a negation.What has the world come to...!
Anyway, back to our story: Lets meet Roombia, named after a...robotic vacuum cleaner! (Okamoto et al. 2009 PLoS ONE)

(Okamoto et al. 2009 PLoS ONE - Roombia truncata; scalebar - 5um)
Sometimes a new organism is found that straightens out and stabilises messy trees (or utterly wrecks well-accepted ones...) The monophyly of our 'Chraptophytes' wasn't completely established until fairly recently, and the relationships of the organisms within that group were also murky. Sometimes the sequences just don't like you and spew out trees that make no sense.

If you go back up to this diagram, the relevance of this finding to the Chromalv. hypothesis is evident in the bottom two trees: the placement of some non-photosynthetic lineages among both of two distinct clades - one with haptos and one with cryptos - takes care of one more troublesome clade with long stretches of basal non-photosynthetic crap chomping away at the likelihood of a single origin. However, there's still the issue of whether Hacrobia is sister to Stramenopiles + Alveolates (good for Chromalv.) or Archaeplastids (bad); as well as establishing whether the "stramalveolate" (can we call it that for now?) plastids and Hacrobia plastids are likely to share a common origin between them.
So it looks like the chromalveolate hypothesis is doing a little bit better, now that not all the basal lineages are non-photosynthetic; multiple plastid loss seems more plausible. However, if you look at one of the broader trees in the paper, it seems to show a pretty well-supported (bootstrap of 0.98) branching of Crhaptos with Archaeplastida. That would be very bad (eg. . However, this is only one gene, Hsp90, and the paper doesn't really discuss that anywhere. Furthermore, my sources inform me that the root(Archaeplastids, Crhapto, (Rhiz, Stramenopiles, Alveolates)) branching seems more likely at the moment. In fact, the little barely-noticeable hyphens under the branchings mean that less that half of the trees support that topology, so ignore everything I just said about archaeplastids and crhaptos. This is to remind us that you can't infer anything from looking at a single tree - biological data is inherently messy, no matter how pretty and crisp and clean the diagram...
By the way, this is by far not all there is to work around the Chromalveolate Hypothesis. There's so much more data pointing in both directions, but I have neither the time nor the qualification to write about it all... and hopefully I haven't butchered too much stuff in this post.
I guess the main thing to take home from all this is how messy and complicated the task of establishing relationships between organisms can be. One must be very skeptical of any models based on single evidence, such as single gene trees or morphology. Or even multiple gene trees, if other data is ignored. Weird shit happens in evolution, and some mysterious event in the past can seriously screw up our perception of the present. As can yet-to-be-discovered characters and phenomena: the Archaezoa were doing quite well until most of the 'amitochondriate' lineages were found to actually contain relics of mitochondrial presence. New data can completely turn trees upside down, roots left dangling all over the place. New data can also strengthen a model. You just never really know, and have to rely on logic, likelihoods and, admittedly, a fair amount of hunches. One must be flexible in the face of new evidence, yet skeptical of the interpretations and validity thereof. Such is life in the fragile wedlock of skepticism and wonder that is science.
PS: Image searches can lead to strange places. Like CreationWiki. Owww. I can feel parts of my brain rotting away and draining out of various cranial orifices:
"All haptophytes are asexual" WRONG. And WHAT THE FLYING FUCK does Blastocystis have to do with Apicomplexa?! Oh right, we reject evolution, therefore phylogeny no longer matters, you polyphylophilic* ignorant morons. Note the absense of references: everything seems to be stolen from Wikipedia et al. Shocking. AND OH MY FSM: "Baraminology"! o_O I'll just leave it at that...
*Adj. Having a fetish for knowingly using polyphyletic lineages. A known side effect of phylogenetic ignorance. Actually, I feel kinda bad bullying the 'epistemologically challenged'. It's more fun to pick on bad PNAS papers instead...
PPS: I'm a nerd: [after subscribing to Trends Ecol Evol feed via Reader]"I've singlehandedly just raised TrEE's IF by a good 10 points! "
My friend (hi!=D) is just evil: "send there a paper to pull it back."
[followed by more impact factor jokes...]
EDIT: David and Jan, thanks for the corrections!
Cavalier-Smith, T. (2002). Chloroplast Evolution: Secondary Symbiogenesis and Multiple Losses Current Biology, 12 (2) DOI: 10.1016/S0960-9822(01)00675-3
KEELING, P., BURGER, G., DURNFORD, D., LANG, B., LEE, R., PEARLMAN, R., ROGER, A., & GRAY, M. (2005). The tree of eukaryotes Trends in Ecology & Evolution, 20 (12), 670-676 DOI: 10.1016/j.tree.2005.09.005
KEELING, P. (2009). Chromalveolates and the Evolution of Plastids by Secondary Endosymbiosis Journal of Eukaryotic Microbiology, 56 (1), 1-8 DOI: 10.1111/j.1550-7408.2008.00371.x
Moore, R., Oborník, M., Janouškovec, J., Chrudimský, T., Vancová, M., Green, D., Wright, S., Davies, N., Bolch, C., Heimann, K., Šlapeta, J., Hoegh-Guldberg, O., Logsdon, J., & Carter, D. (2008). A photosynthetic alveolate closely related to apicomplexan parasites Nature, 451 (7181), 959-963 DOI: 10.1038/nature06635
Okamoto, N., Chantangsi, C., Horák, A., Leander, B., & Keeling, P. (2009). Molecular Phylogeny and Description of the Novel Katablepharid Roombia truncata gen. et sp. nov., and Establishment of the Hacrobia Taxon nov PLoS ONE, 4 (9) DOI: 10.1371/journal.pone.0007080
REYESPRIETO, A., MOUSTAFA, A., & BHATTACHARYA, D. (2008). Multiple Genes of Apparent Algal Origin Suggest Ciliates May Once Have Been Photosynthetic Current Biology, 18 (13), 956-962 DOI: 10.1016/j.cub.2008.05.042
Wow. The discovery of Chromera had completely passed me by.
ReplyDeletemyxotrophy
Mixotrophy. Myxotrophy would probably mean "living off slime"... i and y were not pronounced the same in Classical Greek.
Also, katablepharids.
3am is not a very good time for spelling, but thank you!
ReplyDeleteI think I'm mixing up mixotrophy with myzocytosis, in spelling.
Okamoto et al. 2009 spelled 'opisthokonts' in a rather strange way: 'opistokontes' (see the big tree). They have weird spelling too... =P
I love how you 'animate' the tiny critters by infusing them with your passion! Also your take on the reductionist vs holistic view (which I regard as symbiotic). Great inspiration for an animator. Thank you!
ReplyDeleteI am currently studying this material as an undergrad and surfing the net for some clarity led me to this awesome outlook.
ReplyDeletethankQ for taking the time to write this because I get confused with the protist taxonomy (it doesn't make sense sometimes!)
So now I know to suck it up and remember what I need to know for the exam, and still know that I can trust myself that there must be a better way to organize this tree of organisms. Will I be the one doing it? Who knows, I wish you well and admire your critical outlook.
Cheers!