While reading and writing about neutral evolution, the theme of polarisation in science (pitting of ideas against each other) comes up a lot. Coincidentally, today I had a discussion with someone about group selection and the tendency of issues in academia (and elsewhere in life) to get grotesquely hyperpolarised. A frequent tactic used perhaps almost subconsciously is to pick the most extreme nutcase example from the side you argue against, and use it to represent the entire subfield. Also known as strawman arguments.
This is done almost ubiquitously in academia, and is often not even recognised as a common phenomenon. Perhaps it's some effect of our tribalistic tendencies, I don't know. Government politics is also quite similar. But at least one could be better informed about how their discipline works to be able to correct for some of it on the individual level. I think these practical aspects of philosophy and sociology of science (real fields, btw -- there are people out there who go into science labs and study labrats...very meta.) should be taught along with basic stats and hypothesis testing. Knowing how your field -- and academia in general -- works is arguably more important than memorising Rho GTPase associated pathways.
Curiously, perhaps some of the vicious arguments in science, such as group-level vs. individual-level selection, are actually similar to arguing over the superiority of cartesian vs. polar coordinates in math, rather than using the optimal one for the given circumstances. That is, if you have a flat plane to model, you'd probably go with cartesian coordinates where it's very easy to describe; defining a plane by polar coordinates is cumbersome and absurd (in most cases). Conversely, describing spherical objects would probably favour polar coordinates which simply work better for that type of thing. Similarly, depending on what you're trying to model, you can pick whether group selection or individual selection best fits the situation. Just like both polar and cartesian coordinates exist in the same reality and are ultimately interconvertible, group and individual selection are also both extant and suitable for different situations. The war is utterly unnecessary.
I like that analogy so much I'll breech my own policy of avoiding exposing who I happen to interact with and attribute it to its author -- Wayne Maddison. Now I wonder how many other raging wars in evolutionary theory and beyond are actually arguing over the existence of polar vs. cartesian coordinate systems... selectionism vs. neutralism is one such area that comes to mind.
People in science seriously need to stop obsessing over having a single all-encompassing model for everything, especially in messy fields like biology (fields that deal with real world data, that is). Theoretically, it all does boil down to quantum physics+relativity (or whatever the hypothetical unified version of that would be), but almost any biologist will tell you we actually could care less. Some models are more practical in given situations than others. That doesn't mean one is absolutely superior to another. In order to get anyone, one has to be rather pragmatic. Unless there are actually people in science who think our business is dealing with The Truth. I sincerely hope only undergrads can be that deluded...
PS: Neutral evol part II on its way, not tonight though: really tired and test tomorrow =(

In defense of constructive neutral evolution - Part I
Caution: What follows is mostly an opinion piece by an undergrad. While said undergrad has done a fair amount of reading on the topic, the post is still subject to many errors. Tread carefully. [/disclaimer]
 I won't go into an all-out discussion of neutral evolution here: I'm neither qualified enough nor have enough spare time at the moment. However, some issues seem to crop up multiple times, both here and on other blogs. I figured I'd try to briefly adress some of them, although do take my discussion with a grain of salt. That said, while neutral theory require a certain amount of effort to grasp properly (just like any other aspect of evolutionary biology), it is not something worth dismissing. In fact, if you consider how horribly misunderstood evolutionary biology is on the whole (even among grad students: Gregory & Ellis 2008 BioScience), the neutral elements of evolution seem to be understood by a very small fraction of biologists even.
I won't go into an all-out discussion of neutral evolution here: I'm neither qualified enough nor have enough spare time at the moment. However, some issues seem to crop up multiple times, both here and on other blogs. I figured I'd try to briefly adress some of them, although do take my discussion with a grain of salt. That said, while neutral theory require a certain amount of effort to grasp properly (just like any other aspect of evolutionary biology), it is not something worth dismissing. In fact, if you consider how horribly misunderstood evolutionary biology is on the whole (even among grad students: Gregory & Ellis 2008 BioScience), the neutral elements of evolution seem to be understood by a very small fraction of biologists even.
Perhaps part of the problem is that adaptive evolution is just...flashier. It makes for fun and fairly simple stories: Eg. the peacock has a huge tail to signal to the females that it has nice genes that would compensate for the problems it causes. This "Good Genes" theory is actually taught in first year curricula, and makes very little sense upon further examination, and definitely does not survive Occam's Razor. A simpler explanation would, of course, entail something like runaway sexual selection (ie. female happened to prefer flashy tail, males with flashier tails outcompete their dimmer counterparts, tail gets longer) or that the tail may have a more important function, like scaring away predators. In any case, the adaptive approach very often results in what is mostly a story-telling exercise, and one that is very difficult to deal with experimentally. Worst of all, those stories very easily make sense upon first glance, and thus the field becomes cluttered with poorly thought out theories that sound reasonable.
This post became way too long, so I broke it up into three parts; table of contents here:
Part I
-Adaptationism vs. Neutralism
-"Population genetics ignores reality!"
-Existence of neutrality and near-neutrality
Part II
-Neutral evolution is relevant
-Evolution lacks foresight; it can neither anticipate nor respond
-Rise of complexity through non-adaptive means
-Further examples of constructive neutral evolution
Part III
-Discussion of what sparked this argument: Evolution of ciliate nuclear dimorphism
Adaptationism vs. Neutralism
Of course, none of what I said is new by any margin: the famous Gould and Lewontin 1979 Spandrels of San Marco (free access) paper does a nice job at pointing out many of the fallbacks of panadaptationism. And I don't find it much of an 'attack', as it is often described by diehard adaptationists, but perhaps that's just me. Since so many before me have pointed out the fallbacks of the 'adaptationist programme', I won't bother dwelling on it any further. Besides, this type of thing causes a great polarising effect on the community, with people being either hardcore adaptationists or hardcore neutralists. This seriously fucks up any progress on the topic, because biology hardly tolerates dichotomies. In fact, the truth in this case does not even lie 'somewhere in the middle', but in the fact that both adaptive and neutral processes work in tandem.
Let me reiterate that: Selective and neutral mechanisms work in tandem. Simultaneously. In some situations (eg. large effective population sizes, in bacteria; Lynch 2007 PNAS, Yi 2005 Bioessays), adaptive processes are more dominant; in some cases, adaptive 'forces' are negligible compared to neutral phenomena (small effective population sizes). Considering some specific structure, parts of its evolutionary history have been driven more by drift and mutational bias, interspersed with parts dominated by selective pressures. It's not like selection takes a nap for a while, and then gets back to work while drift, bias et al. chill out. There is no point to dismiss one or the other, like so many tend to do.
Curiously, I've heard numerous times that "Well, maybe selection is less important for bacteria, but it is the dominant force in vertebrates". How hilarious is it that any population geneticist will tell you the exact opposite: bacteria are under overwhelming selective pressure due to their freaking massive effective pop sizes, meaning that drift is much less effective in that situation relative to selection. Vertebrates are actually an awesome example of selection going rather easy: being a large multicellular thing with a backbone is a damn stupid way to copy your genes. Seriously!
Now, you may wonder why should anyone who's not an evolutionary biologist care about any of this? For a cell or developmental biologist, why not assume everything is there for a reason?
Because this leads to rather convoluted explanations for things. Take signalling pathways, for example. Is there any particular reason you have tens of genes required to turn on a particular behaviour in the end when you could've 'designed' it instead to use only one or two? Here we have again a problem with the adaptationist approach: you can pretty much always think of some reason why something is 'useful' or adaptive. That doesn't make it right. What if some features of these pathways originally evolved as a form of adaptively-neutral 'bloating' of the system? See Lynch 2007 PNAS, Lynch 2007 Nature Rev Genet for more on non-adaptive processes in evolution of genetic pathways. (Or just stop reading this post and go through this short list instead ;-)).
"Population genetics ignores reality!"
Now, there's some complaints that popgen kind of fails at taking reality into account. For some work in the field, that is true -- just like in any other field. Did you seriously think everyone, to the last moron, is in touch with reality in your field? If so, I'd love to hear! (no philosophers need apply, heh... although to be fair, there are some 'fringe lunatics' there who actually make sense by our standards.) That said, mathematical modeling requires simplifications to be made to get somewhere. If you have a problem with that, note all those humanities scholars who are sneering at us because we make simplifications as scientists! They wisely take the easy way out and conclude that reality is a social construct and understanding is actually impossible and thus not worthy losing sleep over...
Good mathematical biologists note their simplifications, keep track of them, and know when to simplify what, and what the limitations of their models are. Even better mathematical biologists test their models empirically, thereby producing work that is truly relevant to the rest of us. In my [admittedly still quite inexperienced] opinion, Michael Lynch belongs to that category. Seriously, I despised and dismissed the entire field of population genetics as well, and thanks to some of his papers realised it's probably not a very good idea to do that. In a field as messy as biology, any tidbit of information, even if it comes from simulations, is not only valuable, but essential to the very hope of sorting stuff out. We, that is -- all biologists -- are in no position to discard entire fields because of our petty tribalistic snobbery. Tread with caution -- yes. Dismiss without a second thought -- absolutely not.
Existence of neutrality and near-neutrality
To touch on the existence of 'true' neutrality, let me show you a few diagrams. The first one outlines the history of selectionist and neutral theories:

Story of neutralism and selectionism. (Bernardi 2007 PNAS, OA) Similar diagram and accompanying story can also be found in Ohta 2002 PNAS
You can also see Tomoko Ohta's big review here: Ohta 1992 Annu Rev Ecol Syst (free access).
The major breakthrough here is not even so much the Neutral Theory (which was more of a wake-up call to the stagnating ultra-selectionists), but the Nearly Neutral Theory. Accepting near-neutrality enables one to deal with truly messy and ambiguous situations that don't fit so neatly into the advantageous-deleterious dichotomy. Furthermore, it allows a full spectrum, encorporating the fact that some deleterious mutations are worse than others, and vice versa for the advantageous ones. The 'strict-ness' of selection is further affected by additional factors, such as effective population size.
In short, selection acts probabilistically, not absolutely:

The probability of selection-dependent fixation of an allele vs. product of effective population size(Ne) and selection coefficient(s) of the allele. Note that holding s constant and increasing Ne shifts the selection-dependent fixation probability away from 1 (neutral). Thus, selection is probabilistic; there is also a time dependency here: the longer you run a given situation, the greater the chance that even the slightest deviation from neutrality will be acted on by selection. Given either infinite time or infinite population size, we would see a sharp cut off between advantageous and deleterious alleles. Neutrality would disappear. However, given that no biological system persists for infinite time or has an infinite population size, what one gets is a zone of effectively neutral (Nearly Neutral) mutations; again, the width of this 'zone' is inversely proportional to Ne Disclaimer: I am not well-versed in popgen by any means, please correct me where I'm wrong! (Yi 2006 BioEssays)
The further a mutation deviates in either direction from the true neutral (here assumed to be the same as previous state of the allele, in the context of mutations), the more likely selection is to do something about it, wither selecting against the allele in question, or selecting against those who lack the allele in question (aka "positive selection"). Crudely put, the larger the effective population size, the steeper the selection curve (ie slightly deleterious now becomes quite deleterious, etc).
This actually means that according to pop size alone, selection is far stricter in bacteria than it is in vertebrates. This makes sense: bacteria exhibit a lot less 'design stupidity', if you will, than we do. Again, creating a multicellular organism with a ridiculously long and expensive generation span and a pathetically low reproduction rate is about the stupidest way to perpetrate a handful of genes. But it is tolerated -- in large part because there is a niche for it beyond the reach of more efficient organisms, and to some extent because the effective population size there is so small as to allow the system to drift towards foolish complexity.
Want concrete examples? Genome complexity is a really nice one; I won't discuss details here, but see Yi 2006, Lynch 2007, Stoltzfus 1999, and more Lynch if interested.
To make take an analogy from ecology, a similar argument can be made for the existence of commensalism. Yes, strictly speaking, if you measure all effects and interactions to the finest detail, there are no commensal relationships -- even the slighest extra drag produced by free-riding fish on the manta ray (provided they don't do anything beneficial to it, too lazy to research) would be harmful thereby rendering the relationship parasitic. However, just as in the case of selection, there lies a fuzzy line between parasitism and mutualism, in many cases fuzzy enough to contain 'nearly-parasitic' and 'nearly-commensal' interactions. In that case, the relationship would not be harmful or beneficial enough to really matter within a finite (and turbulent) timeframe. Thus, it is probably more useful to use 'commensal' (well, nearly-commensal) than have to always scratch your head over whether the net total of a relationship is mutualistic or parasitic.
Now that we are hopefully at least considering the possibility of neutral mutations, why should anyone care? Aren't pure selectionists just a strawman anyway? Besides, aren't neutral mutations incapable of really doing anything useful or noticeable? And what the hell is constructive neutral evolution anyway? Stay tuned for this and more in Part II!
References and further reading:
Bernardi, G. (2007). The neoselectionist theory of genome evolution Proceedings of the National Academy of Sciences, 104 (20), 8385-8390 DOI: 10.1073/pnas.0701652104
Gregory, T., & Ellis, C. (2009). Conceptions of Evolution among Science Graduate Students BioScience, 59 (9), 792-799 DOI: 10.1525/bio.2009.59.9.11
Lynch, M. (2007). Colloquium Papers: The frailty of adaptive hypotheses for the origins of organismal complexity Proceedings of the National Academy of Sciences, 104 (suppl_1), 8597-8604 DOI: 10.1073/pnas.0702207104
Lynch, M. (2007). The evolution of genetic networks by non-adaptive processes Nature Reviews Genetics, 8 (10), 803-813 DOI: 10.1038/nrg2192
Ohta, T. (1992). The Nearly Neutral Theory of Molecular Evolution Annual Review of Ecology and Systematics, 23 (1), 263-286 DOI: 10.1146/annurev.ecolsys.23.1.263
Ohta, T. (2002). Inaugural Article: Near-neutrality in evolution of genes and gene regulation Proceedings of the National Academy of Sciences, 99 (25), 16134-16137 DOI: 10.1073/pnas.252626899
Stoltzfus A (1999). On the possibility of constructive neutral evolution. Journal of molecular evolution, 49 (2), 169-81 PMID: 10441669
Yi, S. (2006). Non-adaptive evolution of genome complexity BioEssays, 28 (10), 979-982 DOI: 10.1002/bies.20478
Perhaps part of the problem is that adaptive evolution is just...flashier. It makes for fun and fairly simple stories: Eg. the peacock has a huge tail to signal to the females that it has nice genes that would compensate for the problems it causes. This "Good Genes" theory is actually taught in first year curricula, and makes very little sense upon further examination, and definitely does not survive Occam's Razor. A simpler explanation would, of course, entail something like runaway sexual selection (ie. female happened to prefer flashy tail, males with flashier tails outcompete their dimmer counterparts, tail gets longer) or that the tail may have a more important function, like scaring away predators. In any case, the adaptive approach very often results in what is mostly a story-telling exercise, and one that is very difficult to deal with experimentally. Worst of all, those stories very easily make sense upon first glance, and thus the field becomes cluttered with poorly thought out theories that sound reasonable.
This post became way too long, so I broke it up into three parts; table of contents here:
Part I
-Adaptationism vs. Neutralism
-"Population genetics ignores reality!"
-Existence of neutrality and near-neutrality
Part II
-Neutral evolution is relevant
-Evolution lacks foresight; it can neither anticipate nor respond
-Rise of complexity through non-adaptive means
-Further examples of constructive neutral evolution
Part III
-Discussion of what sparked this argument: Evolution of ciliate nuclear dimorphism
Adaptationism vs. Neutralism
Of course, none of what I said is new by any margin: the famous Gould and Lewontin 1979 Spandrels of San Marco (free access) paper does a nice job at pointing out many of the fallbacks of panadaptationism. And I don't find it much of an 'attack', as it is often described by diehard adaptationists, but perhaps that's just me. Since so many before me have pointed out the fallbacks of the 'adaptationist programme', I won't bother dwelling on it any further. Besides, this type of thing causes a great polarising effect on the community, with people being either hardcore adaptationists or hardcore neutralists. This seriously fucks up any progress on the topic, because biology hardly tolerates dichotomies. In fact, the truth in this case does not even lie 'somewhere in the middle', but in the fact that both adaptive and neutral processes work in tandem.
Let me reiterate that: Selective and neutral mechanisms work in tandem. Simultaneously. In some situations (eg. large effective population sizes, in bacteria; Lynch 2007 PNAS, Yi 2005 Bioessays), adaptive processes are more dominant; in some cases, adaptive 'forces' are negligible compared to neutral phenomena (small effective population sizes). Considering some specific structure, parts of its evolutionary history have been driven more by drift and mutational bias, interspersed with parts dominated by selective pressures. It's not like selection takes a nap for a while, and then gets back to work while drift, bias et al. chill out. There is no point to dismiss one or the other, like so many tend to do.
Curiously, I've heard numerous times that "Well, maybe selection is less important for bacteria, but it is the dominant force in vertebrates". How hilarious is it that any population geneticist will tell you the exact opposite: bacteria are under overwhelming selective pressure due to their freaking massive effective pop sizes, meaning that drift is much less effective in that situation relative to selection. Vertebrates are actually an awesome example of selection going rather easy: being a large multicellular thing with a backbone is a damn stupid way to copy your genes. Seriously!
Now, you may wonder why should anyone who's not an evolutionary biologist care about any of this? For a cell or developmental biologist, why not assume everything is there for a reason?
Because this leads to rather convoluted explanations for things. Take signalling pathways, for example. Is there any particular reason you have tens of genes required to turn on a particular behaviour in the end when you could've 'designed' it instead to use only one or two? Here we have again a problem with the adaptationist approach: you can pretty much always think of some reason why something is 'useful' or adaptive. That doesn't make it right. What if some features of these pathways originally evolved as a form of adaptively-neutral 'bloating' of the system? See Lynch 2007 PNAS, Lynch 2007 Nature Rev Genet for more on non-adaptive processes in evolution of genetic pathways. (Or just stop reading this post and go through this short list instead ;-)).
"Population genetics ignores reality!"
Now, there's some complaints that popgen kind of fails at taking reality into account. For some work in the field, that is true -- just like in any other field. Did you seriously think everyone, to the last moron, is in touch with reality in your field? If so, I'd love to hear! (no philosophers need apply, heh... although to be fair, there are some 'fringe lunatics' there who actually make sense by our standards.) That said, mathematical modeling requires simplifications to be made to get somewhere. If you have a problem with that, note all those humanities scholars who are sneering at us because we make simplifications as scientists! They wisely take the easy way out and conclude that reality is a social construct and understanding is actually impossible and thus not worthy losing sleep over...
Good mathematical biologists note their simplifications, keep track of them, and know when to simplify what, and what the limitations of their models are. Even better mathematical biologists test their models empirically, thereby producing work that is truly relevant to the rest of us. In my [admittedly still quite inexperienced] opinion, Michael Lynch belongs to that category. Seriously, I despised and dismissed the entire field of population genetics as well, and thanks to some of his papers realised it's probably not a very good idea to do that. In a field as messy as biology, any tidbit of information, even if it comes from simulations, is not only valuable, but essential to the very hope of sorting stuff out. We, that is -- all biologists -- are in no position to discard entire fields because of our petty tribalistic snobbery. Tread with caution -- yes. Dismiss without a second thought -- absolutely not.
Existence of neutrality and near-neutrality
To touch on the existence of 'true' neutrality, let me show you a few diagrams. The first one outlines the history of selectionist and neutral theories:

Story of neutralism and selectionism. (Bernardi 2007 PNAS, OA) Similar diagram and accompanying story can also be found in Ohta 2002 PNAS
You can also see Tomoko Ohta's big review here: Ohta 1992 Annu Rev Ecol Syst (free access).
The major breakthrough here is not even so much the Neutral Theory (which was more of a wake-up call to the stagnating ultra-selectionists), but the Nearly Neutral Theory. Accepting near-neutrality enables one to deal with truly messy and ambiguous situations that don't fit so neatly into the advantageous-deleterious dichotomy. Furthermore, it allows a full spectrum, encorporating the fact that some deleterious mutations are worse than others, and vice versa for the advantageous ones. The 'strict-ness' of selection is further affected by additional factors, such as effective population size.
In short, selection acts probabilistically, not absolutely:
The probability of selection-dependent fixation of an allele vs. product of effective population size(Ne) and selection coefficient(s) of the allele. Note that holding s constant and increasing Ne shifts the selection-dependent fixation probability away from 1 (neutral). Thus, selection is probabilistic; there is also a time dependency here: the longer you run a given situation, the greater the chance that even the slightest deviation from neutrality will be acted on by selection. Given either infinite time or infinite population size, we would see a sharp cut off between advantageous and deleterious alleles. Neutrality would disappear. However, given that no biological system persists for infinite time or has an infinite population size, what one gets is a zone of effectively neutral (Nearly Neutral) mutations; again, the width of this 'zone' is inversely proportional to Ne Disclaimer: I am not well-versed in popgen by any means, please correct me where I'm wrong! (Yi 2006 BioEssays)
The further a mutation deviates in either direction from the true neutral (here assumed to be the same as previous state of the allele, in the context of mutations), the more likely selection is to do something about it, wither selecting against the allele in question, or selecting against those who lack the allele in question (aka "positive selection"). Crudely put, the larger the effective population size, the steeper the selection curve (ie slightly deleterious now becomes quite deleterious, etc).
This actually means that according to pop size alone, selection is far stricter in bacteria than it is in vertebrates. This makes sense: bacteria exhibit a lot less 'design stupidity', if you will, than we do. Again, creating a multicellular organism with a ridiculously long and expensive generation span and a pathetically low reproduction rate is about the stupidest way to perpetrate a handful of genes. But it is tolerated -- in large part because there is a niche for it beyond the reach of more efficient organisms, and to some extent because the effective population size there is so small as to allow the system to drift towards foolish complexity.
Want concrete examples? Genome complexity is a really nice one; I won't discuss details here, but see Yi 2006, Lynch 2007, Stoltzfus 1999, and more Lynch if interested.
To make take an analogy from ecology, a similar argument can be made for the existence of commensalism. Yes, strictly speaking, if you measure all effects and interactions to the finest detail, there are no commensal relationships -- even the slighest extra drag produced by free-riding fish on the manta ray (provided they don't do anything beneficial to it, too lazy to research) would be harmful thereby rendering the relationship parasitic. However, just as in the case of selection, there lies a fuzzy line between parasitism and mutualism, in many cases fuzzy enough to contain 'nearly-parasitic' and 'nearly-commensal' interactions. In that case, the relationship would not be harmful or beneficial enough to really matter within a finite (and turbulent) timeframe. Thus, it is probably more useful to use 'commensal' (well, nearly-commensal) than have to always scratch your head over whether the net total of a relationship is mutualistic or parasitic.
Now that we are hopefully at least considering the possibility of neutral mutations, why should anyone care? Aren't pure selectionists just a strawman anyway? Besides, aren't neutral mutations incapable of really doing anything useful or noticeable? And what the hell is constructive neutral evolution anyway? Stay tuned for this and more in Part II!
References and further reading:
Bernardi, G. (2007). The neoselectionist theory of genome evolution Proceedings of the National Academy of Sciences, 104 (20), 8385-8390 DOI: 10.1073/pnas.0701652104
Gregory, T., & Ellis, C. (2009). Conceptions of Evolution among Science Graduate Students BioScience, 59 (9), 792-799 DOI: 10.1525/bio.2009.59.9.11
Lynch, M. (2007). Colloquium Papers: The frailty of adaptive hypotheses for the origins of organismal complexity Proceedings of the National Academy of Sciences, 104 (suppl_1), 8597-8604 DOI: 10.1073/pnas.0702207104
Lynch, M. (2007). The evolution of genetic networks by non-adaptive processes Nature Reviews Genetics, 8 (10), 803-813 DOI: 10.1038/nrg2192
Ohta, T. (1992). The Nearly Neutral Theory of Molecular Evolution Annual Review of Ecology and Systematics, 23 (1), 263-286 DOI: 10.1146/annurev.ecolsys.23.1.263
Ohta, T. (2002). Inaugural Article: Near-neutrality in evolution of genes and gene regulation Proceedings of the National Academy of Sciences, 99 (25), 16134-16137 DOI: 10.1073/pnas.252626899
Stoltzfus A (1999). On the possibility of constructive neutral evolution. Journal of molecular evolution, 49 (2), 169-81 PMID: 10441669
Yi, S. (2006). Non-adaptive evolution of genome complexity BioEssays, 28 (10), 979-982 DOI: 10.1002/bies.20478
MM17 Answer - Spironucleus: double cells with twisted nuclei

SEMs of diplomonad fish parasite Spironucleus vortens. cr - compound lateral ridge.lpr + rpr - left and right peripheral ridge, respectively. Note their rope-like form. pp - posterior papillum. Note flagellar pockets in (4). r - recurrent flagellum. 5 - an atypical specimen with transposed posterior structures. 6 - laterla view of posterior end. A fairly complicated flagellate! (Sterud & Poynton 2002 JEM)
To clarify the complicated morphology:

Drawings of the 'double-celled' Spironucleus vortens. The two recurrent flagella pass inside the cell and emerge at the posterior end. (Poynton et al. 1995 JEM)
Spironucleus may strike you as being particularly symmetrical. In fact, it very well is a 'double cell', containing two nuclei (slightly wrapping around each other helically, hence Spironucleus), and two sets of flagella. The path of the recurrent flagella makes sense when considering that the single cells of the group have three flagella pointing one way, and the fourth pointing another. These double cells are case of the two cells being 'stuck together' at the side of the recurrent flagellum. Here's a TEM to show the elongate nuclei and the flagella passing between them:

Top: longitudinal section at the anterior end of the flagellate. n - nuclei; k - kinetosomes. Bottom: cross-section of the anterior end. Note the recurrent flagella (r; circled in red) passing between the nuclei. (Poynton et al. 1995 JEM)
Here's another species of Spironucleus, S.berkhanaus, in arctic char blood:

Parasitic Spironucleus barkhanus in the blood of arctic char, as well as in isolation. Note that it's a different species from the one above, which may explain the lack of lateral ridges.(Sterud et al. 2003 Dis Aquatic Organisms)
Note how the two species seem a bit different from each other. This shows the morphological diversity in the group. In fact, Spironucleus seems to be a bit polyphyletic or paraphyletic at best (Jørgensen & Sterud 2007 Protist; Kolisko et al. 2008 BMC Evol Biol).

Phylogeny of Spironucleus. This poor genus seem to be ruthlessly strewn all over Diplomonadida. Note position of S.vortens and S.berkhanus. (Jørgensen & Sterud 2007 Protist)
Diplomonads have an interesting tale involving cell cycle defects and duplications of the nucleus and flagella, but I'll leave you in suspense for a while. That's a bigger topic, and I should probably introduce our cute friend Giardia first (cute friend in SEM, and horrible foe in the intestine...). Giardia is a independent case of cell 'doubling', and is organised quite differently. Further discussion of diplomonads should happen...eventually. Feel free to nag me about it if you're really interested!
References
JORGENSEN, A., & STERUD, E. (2007). Phylogeny of Spironucleus (Eopharyngia: Diplomonadida: Hexamitinae) Protist, 158 (2), 247-254 DOI: 10.1016/j.protis.2006.12.003
Kolisko, M., Cepicka, I., Hampl, V., Leigh, J., Roger, A., Kulda, J., Simpson, A., & Flegr, J. (2008). Molecular phylogeny of diplomonads and enteromonads based on SSU rRNA, alpha-tubulin and HSP90 genes: Implications for the evolutionary history of the double karyomastigont of diplomonads BMC Evolutionary Biology, 8 (1) DOI: 10.1186/1471-2148-8-205
POYNTON, S., FRASER, W., FRANCIS-FLOYD, R., RUTLEDGE, P., REED, P., & NERAD, T. (1995). Spironucleus vortens N. Sp. from the Freshwater Angelfish Pterophyllum scalare: Morphology and Culture The Journal of Eukaryotic Microbiology, 42 (6), 731-742 DOI: 10.1111/j.1550-7408.1995.tb01625.x
Sterud E, Poppe T, & Bornø G (2003). Intracellular infection with Spironucleus barkhanus (Diplomonadida: Hexamitidae) in farmed Arctic char Salvelinus alpinus. Diseases of aquatic organisms, 56 (2), 155-61 PMID: 14598991
STERUD, E., & POYNTON, S. (2002). Spironucleus vortens (Diplomonadida) in the Ide, Leuciscus idus (L.) (Cyprinidae): a Warm Water Hexamitid Flagellate Found in Northern Europe The Journal of Eukaryotic Microbiology, 49 (2), 137-145 DOI: 10.1111/j.1550-7408.2002.tb00357.x
Sunday Protist -- Trichotokara nothriae: Guitar-shaped gregarine
Gregarines are a group of apicomplexans (='Sporozoa', a vastly diverse group famous for the malarial parasite Plasmodium and the behaviour-altering Toxoplasma) characterised by a monoxenous (single host) lifestyle that is quite different from that of other 'apis'. Christopher Taylor wrote a nice post about them here.
Apicomplexa are alveolates along with ciliates and dinoflagellates; you can find them on the left side of this tree . The apicomplexan phylogeny is a complete mess at the moment; the old coccidian-haematozoan-gregarine divisions aren't too well-supported and the relationships of stuff within them are even murkier. As an aside, many apis have an 'apicoplast', or a relic plastid of red algal origin -- their ancestors were once photosynthetic! In fact, a paraphyletic group of organisms basal to apicomplexa (Chromera et al.) are currently photosynthetic, further supporting the photosynthetic ancestry of these weird mostly-intracellular parasites, most of whom rarely ever see the light of day!
Gregarines are typically invertebrate parasites and unlike other apicomplexans, tend to spend most of their lives extracellularly; in fact, their cellular penetration consists of attaching themselves to a cell via the mucron (holdfast-like structure). You can read more about their biology and life cycle on their ToLWeb page. (also a review in Tr Parasitol: Leander 2007) If you want to see some for yourself, kidnapping and slicing up an earthworm is an easy way to do so: Monocystis is a parasite of earthworm seminal vesicles (feeds on sperm), and a rather abundant one. It may actually be quite easy to find various apicomplexan parasites in insects -- it is estimated that most of them may have an api specialised in parasitising them, which hints at the total apicomplexan diversity being something outrageously vast! Such a project would also be a good excuse to learn insect anatomy, which I find to be quite complicated.
Right, you wanted to see a new genus of guitar-shaped gregarines:

Trichotokara from the intestine of an onuphid tubeworm. a-e: trophozoites (feeding forms). M - mucron ('holdfast'), CB - cell body. Arrow - junction between mucron and cell body, which can be seen extending further into the mucron in (e; arrowheads). f: gamonts in syzygy, or gregarine sex. Scalebars: a-e 10um; f 25um. (Rueckert & Leander 2010 J Invert Pathol)
By the way, if anyone asks you for a six-letter word in English 'devoid of any vowels', keep 'syzygy' in mind. Technically it does have vowels, as any phonologist would tell you, but most people insist on equating letters with sounds, and y 'is not a vowel'. Regardless, it's still a really awesome word. Syzygy!
More gregarine awesomeness. Note how the cell surface seems to strive for increased surface area, especially in the mucron which gets inserted into a cell:

SEM of Trichotokara. b - close-up of hair-like projections of the mucron. c - junction between mucron and cell body. d - folds along the cell body. Scalebars: a - 10um; b-d - 1um. (Rueckert & Leander 2010 J Invert Pathol)
This gives me an excuse to mention a paper on proximate vs. ultimate convergence by the senior author on the above gregarine paper: Leander 2008 JEM (free access). Among several other examples of ultimate convergence between multicellular and unicellular organisms inhabiting similar environments, gregarines and nematodes are compared in terms of their structural organisation. While nematodes have longitudinal muscles just beneath the elastic epidermis, gregarines have subpellicular bands of longitudinal microtubules running just underneath the elastic cortex (although used differently -- see gliding motility below). Curiously, in both cases the result is a sinusoidal (wiggling) pattern of movement. Additionally, tapeworm and Haplozoon (dinoflagellate) surface morphology are noted to be similar (covered with microvili), for the obvious purpose of increasing surface area. It's probably not much of a stretch to add gregarine surface structure to that list. (see Leander et al. 2003 J Parasitol for more gregarine surfaces) 
Interesting case of structural ultimate convergence between nematodes and gregarines. Purple - bands of muscle and microtubules, respectively. Blue - elastic epidermis and tri-layered cortex, respectively. The three cortical layers consist of the plasma membrane at the very surface, with two alveolar membranes immediately below. (Leander 2008 JEM)
Before we proceed to a digression on apicomplexan motility, oblicatory phylogeny of Trichotokara and relatives. Note its extremely diverged SSU sequence resulting in a hellishly long branch:
Interesting case of structural ultimate convergence between nematodes and gregarines. Purple - bands of muscle and microtubules, respectively. Blue - elastic epidermis and tri-layered cortex, respectively. The three cortical layers consist of the plasma membrane at the very surface, with two alveolar membranes immediately below. (Leander 2008 JEM)
Before we proceed to a digression on apicomplexan motility, oblicatory phylogeny of Trichotokara and relatives. Note its extremely diverged SSU sequence resulting in a hellishly long branch:

ML tree of SSU rDNA sequences. Probably wouldn't trust its exact placement among the gregarines just yet... (Rueckert & Leander 2010 J Invert Pathol)
Apicomplexans are generally aflagellate in their trophic stage (I say 'generally' just in case...) -- their motility is an interesting topic, as they can't exactly extrude pseudopodia either. Nor do they have any spirochaetes doing the work for them as in Mixotricha, nor do they wildly thrash about an internal bundle of microtubules like Saccinobaculus. So how do they do it? Just like pennate diatoms: by gliding motility.
While sharing some basic similarities with diatom gliding, the apicomplexan variant has an unrelated origin and is quite different. One annoying thing (to us) about alveolates is their alveolae, or little membranous sacs just underneath the plasma membrane. In apicomplexan cell biology literature, this is called the Inner Membrane Complex. Prior to explaining why this detail is particularly annoying, first let's go over the crude basics of gliding motility: First, you need something to anchor to the substrate. This material is usually discarded, leaving behind a trail of 'slime', if you will. Then, you need an adaptor protein [complex] that attaches to the anchoring substance and crosses the plasma membrane. This adaptor must have a way of reaching a cytoskeletal element, usually actin via myosins (eg. see Molino & Wetherbee 2008 Biofouling; also, that journal title is very WTF...). The problem (again, mostly for researchers, and students...) with apicomplexa is their tendency to have the Inner Membrane Complex in the middle of that. This means the mechanism looks roughly like this:

{kind=link}
Or, in the language of Nature Reviews:

Cell biology: Always more fun with extra gene/protein names thrown in, especially those irrelevant to the point. Shall we look up some protein structures while we're at it?
Ignore the target cell part -- a similar process happens along other surfaces too. If I recall, the model with intra-IMC proteins reaching across between actin and microtubule systems is actually more up-to-date; the "rolling IMC conveyor belt" model was outdated. Don't quote me on this though! (Baum et al. 2006 Nature Rev Microbiol)
Remember cramming the molecular biology of amoeboid motion? Isn't it almost a good thing that traditional cell biology courses are so phylogenetically impoverished? So many things are much more complex than animal cells, so we actually get the easy (and [arguably] boring) option. In a nutshell, you have something like this:
anchor-adhesin-actin-myosin-[interamembranous particles?]-subpellicular microtubulesThe myosins move to the opposite of cell motility (and actin polymerisation), thereby pushing the pellicular microtubule skeleton in the right direction. Look at the Soldati & Meissner figure again to see why. It's a rather convoluted process just to get a cell moving! Of course, that complexity is more of a problem for cell biologists than the organism, considering how abundant and efficient apicomplexans tend to be.
Another aside: Apicomplexans, as well as numerous other organisms, are capable of a cell divison process known as palintomy: they can undergo several rounds of mitosis without cytokinesis, resulting in multinucleate cells (helps to not have open mitosis), and then simultaneously undergo cytokinesis for each of those nuclei (cellularisation). In gregarines, this looks vaguely like budding, as the nuclei tend to congregate near the cortex during this process (Kuriyama et al. 2005 Cell Motility Cytosk). Drosophila embryos do something similar, so palintomy isn't that unusual, but still pretty cool.
Back to gregarines, there are some more species that seem to be on a morphological acid trip. Some of them have been described only once and never noted again, which makes me sad:

Aikinetocystis singularis. I really want an SEM of that! Too bad it's from an obscure burmese earthworm... (Gates 1926 Biol Bulletin)
So if you like finding new species and genera and describing them, may I recommend apicomplexan diversity. It's taking a while for entrail-hungry parasitologists to go through all the various invertebrate (and vertebrate) parasites out there, so there's still plenty of room for work. If there is truly one species of apicomplexa for roughly each species of insects (and other animals), that pie chart of diversity showing most life as insects (and protists but a tiny splinter) is truly laughable:

LOL. Simply hilarious! Looks like the "global biodiversity assessment" team was a bit short on microbiologists... (at least they admit to not knowing much bacterial diversity; at least they put 'protozoa' and 'algae' in quotation marks...) (Purvis & Hector 2000 Nature)
LOL. Simply hilarious! Looks like the "global biodiversity assessment" team was a bit short on microbiologists... (at least they admit to not knowing much bacterial diversity; at least they put 'protozoa' and 'algae' in quotation marks...) (Purvis & Hector 2000 Nature)
All hail microbial parasites -- the bane of biodiversity research!
References
Baum, J., Papenfuss, A., Baum, B., Speed, T., & Cowman, A. (2006). Regulation of apicomplexan actin-based motility Nature Reviews Microbiology, 4 (8), 621-628 DOI: 10.1038/nrmicro1465
G. E. Gates (1926). Preliminary Note on a New Protozoan Parasite of Earthworms of the Genus Eutyphœus Biological Bulletin, 51 (6), 400-404
LEANDER, B. (2008). Marine gregarines: evolutionary prelude to the apicomplexan radiation? Trends in Parasitology, 24 (2), 60-67 DOI: 10.1016/j.pt.2007.11.005
LEANDER, B. (2008). A Hierarchical View of Convergent Evolution in Microbial Eukaryotes Journal of Eukaryotic Microbiology, 55 (2), 59-68 DOI: 10.1111/j.1550-7408.2008.00308.x
Molino, P., & Wetherbee, R. (2008). The biology of biofouling diatoms and their role in the development of microbial slimes Biofouling, 24 (5), 365-379 DOI: 10.1080/08927010802254583
Purvis, A., & Hector, A. (2000). Getting the measure of biodiversity Nature, 405 (6783), 212-219 DOI: 10.1038/35012221
Rueckert, S., & Leander, B. (2010). Description of Trichotokara nothriae n. gen. et sp. (Apicomplexa, Lecudinidae) – an intestinal gregarine of Nothria conchylega (Polychaeta, Onuphidae) Journal of Invertebrate Pathology DOI: 10.1016/j.jip.2010.03.005
Soldati, D., & Meissner, M. (2004). Toxoplasma as a novel system for motility Current Opinion in Cell Biology, 16 (1), 32-40 DOI: 10.1016/j.ceb.2003.11.013
References
Baum, J., Papenfuss, A., Baum, B., Speed, T., & Cowman, A. (2006). Regulation of apicomplexan actin-based motility Nature Reviews Microbiology, 4 (8), 621-628 DOI: 10.1038/nrmicro1465
G. E. Gates (1926). Preliminary Note on a New Protozoan Parasite of Earthworms of the Genus Eutyphœus Biological Bulletin, 51 (6), 400-404
LEANDER, B. (2008). Marine gregarines: evolutionary prelude to the apicomplexan radiation? Trends in Parasitology, 24 (2), 60-67 DOI: 10.1016/j.pt.2007.11.005
LEANDER, B. (2008). A Hierarchical View of Convergent Evolution in Microbial Eukaryotes Journal of Eukaryotic Microbiology, 55 (2), 59-68 DOI: 10.1111/j.1550-7408.2008.00308.x
Molino, P., & Wetherbee, R. (2008). The biology of biofouling diatoms and their role in the development of microbial slimes Biofouling, 24 (5), 365-379 DOI: 10.1080/08927010802254583
Purvis, A., & Hector, A. (2000). Getting the measure of biodiversity Nature, 405 (6783), 212-219 DOI: 10.1038/35012221
Rueckert, S., & Leander, B. (2010). Description of Trichotokara nothriae n. gen. et sp. (Apicomplexa, Lecudinidae) – an intestinal gregarine of Nothria conchylega (Polychaeta, Onuphidae) Journal of Invertebrate Pathology DOI: 10.1016/j.jip.2010.03.005
Soldati, D., & Meissner, M. (2004). Toxoplasma as a novel system for motility Current Opinion in Cell Biology, 16 (1), 32-40 DOI: 10.1016/j.ceb.2003.11.013
Moss microforay teaser
Really busy lately, hence the spotty blogging as of late. However, tonight I just happened to have a small moss sample and some 'spare time' (reads: time I could've used productively instead...) after imaging some lab-related stuff at the scope. Went on a bit of a moss microforay, found loads of Euglyphids, and will post a couple teasers as it will take me a while to process all the images:


Sadly, most of my Euglyphid images were made before I realised some genius managed to get oil encrusted on the 40x air objective. The subsequent avid cleaning accompanied by profanity did help clear up the lens a little, but it's still seriously fucked. That kind of thing makes me really angry, as it's easily avoidable and keeping lenses clean can improve your image quality so much for free. A clean simple lens can get better images than a dirty fancy lens, and each speck/layer of crap eats away a nice chunk of resolution. Look at the difference using the same specimen (which drifted a little during the distraction) - the one on the left is before cleaning, the one on the right after (the left image is at the optimal DIC settings, for the right image the analyser and prism haven't been properly re-adjusted yet, so it gets better):


The microforay images, presentation part 2 narration, proper response to a certain comment and maybe even a couple other obligations (eg. Mystery Micrograph write-ups...) will be worked on this weekend. In any case, I'll finish the narration then, sorry for keeping you waiting!
Edit 27.03.10: Finished narration for part 2! Enjoy, please feel free to ask questions, etc.



Another thing I noticed was that suddenly there was A LOT MORE LIGHT. So yeah, keep your lenses clean and if the image is strangely fuzzy, don't forget to check if some idiot dirtied the objective somehow. Oh, and keep oil the fucking hell away from any non-oil objectives, it's not that bloody difficult!!!
Grrr I really hate it when so much time gets wasted because of something really stupid being wrong with the scope.
Anyway, there was also a mystery metazoan of some sort. I can't quite figure out what it is. Does anyone know? I'm no good with worm-like metazoa...
Grrr I really hate it when so much time gets wasted because of something really stupid being wrong with the scope.
Anyway, there was also a mystery metazoan of some sort. I can't quite figure out what it is. Does anyone know? I'm no good with worm-like metazoa...

Edit 27.03.10: Finished narration for part 2! Enjoy, please feel free to ask questions, etc.
Sunday Protist -- Aspidisca: Walking ciliates with scrambled genomes
That's right, instead of swimming around like a normal flagellate must do, these creatures have thick bundles of microtubules called cirri which they use as little "feet" to walk on surfaces. One such creature, Aspidisca, apparently loves walking so much it rarely ever swims (Banchetti et al. 2003 Can J Zool)!
If that scalebar were 10cm rather than 10um, Aspidisca would make such a wonderful pet! One can almost imagine what a wonderful feeling it would be to have cirri gently brush against the palm of your hand... (Rosati et al. 1987 Trans Am Microscop Soc)
Aspidisca lives in marine coastal sands, which explains why it doesn't really need to swim much. Like most other ciliates, it's a predator, and probably not one you'd like to encounter if it were of our size. Also, like most, if not all, other ciliates, Aspidisca requires sex to make fresh somatic nuclei (Warning: The following figure contains porn.)
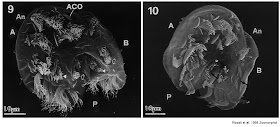
Now, hypotrichs aren't just weird on the surface -- they are also utterly bizzare at heart. To them, having two differentiated nuclei, one resembling a giant bag of linear eukaryotic 'plasmids' and not even doing mitosis properly, is mundane and boring. They go the extra mile and scramble their genomes. That's right, instead of having your regular eukaryotic gene (interspersed with a few introns, of course), they split up fragments of their genes and mix up their order and direction. (This must really scramble the brains of the unfortunate souls forced to do bioinformatics on these organisms!)
Before we go into gene scrambling, a very brief primer on ciliate genomes is in order. As you may already have heard, ciliates have a transcriptionally-silent micronucleus (MIC) and a transcriptionally-active massively polyploid bag of short chromosomes called a macronucleus (MAC). The MAC may have a ridiculous number of copies of a single chromosome, such as 9000 for some chromsomes in Tetrahymena (Yao & Chao 2005 Annu Rev Genet). The splitting of such nuclei during cell division is a topic worthy of a few whole posts, although I must point out that ciliates have rather insane regulation strategies for DNA replication due to the variable gene copy number after each mitosis.
While the MAC sequences originate from the MIC (right after conjugation), they don't actually match their MIC counterparts exactly; in fact, certain portions are deleted along their way from the MIC! Furthermore, whether a portion is retained or not depends on its presence in the old MAC, which is already destroyed by this point. There's a whole epigenetic process of selected sequence deletion that's also a topic for another time (you can read about it here). Essentially, you have MAC-Destined Sequences (MDS) and Internally Eliminated Sequences (IES), which are retained and excised, respectively. They kind of act like exons and introns on a genomic level. Be very careful though, as it is very easy to begin mixing up IESs and introns -- they're just so alike in sound and concept, and yet so different...

This genome-level "intron" situation gets weirder yet: in some species, the expressed gene fragments (MDSs) are not arranged in a sensible order in the MIC! That is, if you have gene fragments 1-2-3-4 normally, you can sometimes get 2-1-3-4 or even 3-1-4-2, or some other permutation thereof. The MAC sequence remains correct: 1-2-3-4 (otherwise, the organism would die!) Furthermore, those MIC fragments can even be reversed (see above figure)! How do we get from a huge mess in the germline MIC to a regular fully functional sequence in the MAC?
Unscrambling is still poorly understood, but it seems that in addition to IES removal, some form of recombination happens along the way. Furthermore, errors in the process are somehow corrected before the MAC matures. For Oxytricha ACTIN1, IESs seem to be removed before recombination and translocations (Möllenbeck et al. 2008 PLoS ONE; OA). As scrambling is slightly easier to grasp than unscrambling, a bit more seems to be understood about its evolution. It looks like gene scrambling is a fairly likely result of a system involving intricate sequence excision mechanisms, alternatively known a genome rearrangements.


Real examples of scrambled genes, and models of their evolution: Left - ACTIN1 in Oxytricha. Shows gene structure and then a model for its unscrambling. x's indicate recombination between paired repeats. Right - Model for initial scrambling of Oxytricha ALPHA-TELOMERE BINDING PROTEIN. Fragments of AT-rich DNA can be randomly inserted into the gene as IESs, to be quietly removed afterwards. By IES insertions when the gene is in some awkward topology (eg. loopy), genes can become scrambled. See source for further reading: Prescott 2000 Nature Rev Genet (free access), as well as Chang et al. 2005 PNAS)
So how do we get from genome rearrangements to scrambling? Enter recombination. If recombination scrambles up pieces of genes in most organisms, they die and we don't really see scrambled genes. However, due to their intricate mechanism of eliminating IESs, ciliates are able to unscramble the gene fragments and arrange them in the correct order. Likewise, they are also able to tolerate random insertions within genes, as they get removed in the somatic nucleus anyway. This excess capacity caused by an effective fixing mechanism enabled ciliates to tolerate disordered gene fragments -- a really nice example of a substantial increase in complexity with hardly any adaptive advantage whatsoever; that is, constructive neutral evolution. The evolution of ciliate gene scrambling is discussed in more detail in Stoltzfus 1999 J Mol Evol.
Strictly speaking, it is not known whether Aspidisca itself engages in this weird gene scrambling mess, as its genome is a mystery. It would probably be useful to check Euplotes -- if it scrambles, than most of the stuff in between it and Oxytricha, Stylonichia, Urostyla et al., including Aspidisca. Regardless, Aspidisca is close enough to the known scramblers, so I did have a right to ramble about it anyway.

In case someone else out there cares where Aspidisca branches in Euplotida. (Shen et al. 2010 Eur J Protistol; AOP)
Ciliates are another great model for both cell and molecular biology. Of the awesome Hypotrichs, Oxytricha has its genome sequenced and is ready to be played with. It also has really awesome morphogenesis stuff, but that's also a topic for a later post.
Almost completely off-topic, but apparently there's protein family specific to Alveolates that colocalises with the surface of the characteristic alveolar sacs just at the cortex: the Alveolins:

Blue - nucleo; Green - alveolin (Gould et al. 2008 MBE; free access)
This is really cool! In vivo flurescence imaging is another one of my obsessions, though immunostaining is ok too, provided it actually works...
References
Banchetti, R., Erra, F., Ricci, N., & Dini, F. (2003). Ethogram of Aspidisca sedigita Canadian Journal of Zoology, 81 (1), 14-20 DOI: 10.1139/z02-194
Gould, S., Tham, W., Cowman, A., McFadden, G., & Waller, R. (2008). Alveolins, a New Family of Cortical Proteins that Define the Protist Infrakingdom Alveolata Molecular Biology and Evolution, 25 (6), 1219-1230 DOI: 10.1093/molbev/msn070
Möllenbeck, M., Zhou, Y., Cavalcanti, A., Jönsson, F., Higgins, B., Chang, W., Juranek, S., Doak, T., Rozenberg, G., Lipps, H., & Landweber, L. (2008). The Pathway to Detangle a Scrambled Gene PLoS ONE, 3 (6) DOI: 10.1371/journal.pone.0002330
Prescott DM (2000). Genome gymnastics: unique modes of DNA evolution and processing in ciliates. Nature reviews. Genetics, 1 (3), 191-8 PMID: 11252748
Rosati, G., Verni, F., & Dini, F. (1998). Mating by conjugation in two species of the genus Aspidisca (Ciliata, Hypotrichida): an electron microscopic study Zoomorphology, 118 (1), 1-12 DOI: 10.1007/s004350050051
Rosati, G., Verni, F., Bracchi, P., & Dini, F. (1987). An Ultrastructural Analysis of the Ciliated Protozoon Aspidisca sp. Transactions of the American Microscopical Society, 106 (1) DOI: 10.2307/3226282
Stoltzfus, A. (1999). On the Possibility of Constructive Neutral Evolution Journal of Molecular Evolution, 49 (2), 169-181 DOI: 10.1007/PL00006540
Shen et al. 2010 European Journal of Protistology (Advance online pub, not yet indexed in DOI or ResearchBlogging)